
The Capsaicin Paradox: Biological Honesty and the Social Poison of the Spicy Food Semiotic Field
The biological honesty of the capsaicin response—the involuntary flush, the beads of sweat, the dilated pupils—is precisely what makes it such fertile ground for social semiotics. Because these reactions are difficult to fake, they are treated as “honest signals” of an internal state. Yet this very treatment reveals a deeper philosophical move: what we call “biological honesty” is itself a form of strategic essentialism—a collective decision to treat the body’s signals as ground truth so that we have a fixed reference point in an otherwise endlessly performative social world. The body’s reactions are not inherently more truthful than any other sign; we designate them as such because a semiotic system requires an anchor, a place where interpretation agrees to stop. This anchoring is not a discovery about nature but a cultural commitment—one that quietly underwrites every social meaning we subsequently extract from the burn.
However, the moment these signals are observed by another, they are pulled into the Semiotic Field, where they are no longer just physiological events but symbols to be interpreted.
The sensation of heat from a chili pepper is a biological absolute—a chemical interaction between capsaicin and the TRPV1 receptors that signals a warning of thermal pain. This is what we might call “biological honesty”: the body reacting to a perceived threat with a predictable, physiological response. However, once this sensation enters the human social sphere, it is immediately transmuted into something far more complex. It becomes a “social poison,” where the raw experience of pain is filtered through layers of cultural meaning, gendered performance, and status signaling. To understand this transformation, we must map the Spicy Food Semiotic Field—a conceptual space where the physiological reality of the burn collides with the symbolic weight of its consumption.
The Biological Honesty of Heat
At its core, the capsaicinoid response is an evolutionary defense mechanism—and a remarkably precise one. Plants of the genus Capsicum developed capsaicin not as a broad-spectrum toxin but as a strategy of Directed Deterrence: a molecular key shaped to fit one specific lock. Capsaicin binds to the TRPV1 vanilloid receptor—a thermosensor calibrated to fire at temperatures above 43°C—and activates it at room temperature. In effect, the molecule lies to the mammalian nervous system, convincing it that the mouth is being scalded when no thermal event is occurring. Birds, lacking the specific TRPV1 configuration sensitive to capsaicin, consume the fruit and disperse the seeds intact through their digestive tracts. The evolutionary elegance is striking: the plant does not poison indiscriminately but engineers a targeted sensory hallucination, exploiting the mammalian pain architecture to redirect its reproductive future through avian vectors. This is the first layer of biological honesty—though, as we shall see, even calling it “honesty” requires qualification, since the signal is, at the molecular level, a sophisticated deception.
When a human bypasses this warning, the resulting sensation acts as a profound embodiment anchor. Unlike the subtle nuances of flavor, the heat of a habanero or a ghost pepper demands immediate, total interoceptive awareness. It forces the consciousness back into the physical frame, demanding attention to the mouth, the throat, and the rising internal temperature. In an era of digital abstraction, this “honest” pain provides a grounding reality that cannot be ignored or intellectualized away. Yet even here, a crucial philosophical nuance intervenes—what we might call the nociceptive-interpretive gap. The TRPV1 receptor performs nociception: a straightforward chemical-to-electrical transduction, converting the presence of capsaicin into an ion channel event and a nerve impulse. But pain—the burning, the panic, the urge to reach for milk—is not the signal itself; it is the brain’s interpretation of that signal, constructed through layers of context, memory, and expectation. Even at the most “biological” level, the body is not simply reporting; it is already narrating. The supposedly raw datum of the burn is, from its very first synapse, an act of meaning-making. This gap between transduction and experience is what makes the entire semiotic apparatus possible: if the signal were truly unmediated, there would be nothing for culture to rewrite.
Physiologically, this heat serves several utilitarian functions. It acts as a built-in appetite throttle, increasing satiety and slowing the pace of consumption—a natural defense against overindulgence. Furthermore, its role as a vasodilator and irritant triggers a sinus-clearing mechanism, facilitating respiratory relief.
The transition from pain to pleasure—the “acquired taste”—is driven by the body’s own compensatory mechanisms. As the TRPV1 receptors signal intense heat, the brain responds by releasing a flood of endorphins and dopamine to mitigate the perceived trauma. This creates the “endorphin loop,” where the initial distress is followed by a mild euphoria. Over time, the brain learns to associate the burn not with damage, but with the subsequent neurochemical reward, transforming a biological warning into a sought-after culinary experience. But there is a deeper physiological process at work beyond mere associative learning: chili bleaching, the functional desensitization of TRPV1-expressing neurons through repeated capsaicin exposure. With chronic stimulation, these neurons undergo a depletion of neuropeptides such as Substance P and, in some cases, a reversible degeneration of their peripheral terminals. The receptor doesn’t “toughen up”; it is effectively silenced. What society reads as “toughness”—the stoic consumption of a Carolina Reaper without visible distress—may simply be the result of bleached receptors, a neurological quiet mistaken for a moral quality. And the body’s encounter with capsaicin does not end at the palate. TRPV1 receptors line the entire gastrointestinal tract, from esophagus to rectum, and capsaicin activates them at every station along the way. This “second burn”—the intestinal cramping, the unmistakable rectal sting hours later—is the plant’s evolutionary parting gift, a final reminder that the directed deterrence strategy extends well beyond the mouth. It is a sensation rarely discussed in polite company, yet it completes the biological circuit: the mammalian body is warned at entry and at exit, bookending the digestive journey with the same molecular lie.
Benign Masochism and the Psychology of the Acquired Taste
Yet the endorphin loop and receptor desensitization, powerful as they are, do not fully explain why humans seek out the burn in the first place. After all, the body produces compensatory endorphins in response to many forms of pain—stubbing a toe, touching a hot stove—without those experiences becoming objects of craving. The missing piece is not neurochemical but cognitive, and it was most precisely articulated by the psychologist Paul Rozin, who coined the term “benign masochism” to describe the peculiar human capacity to derive pleasure from sensations that the body’s alarm systems register as threats. Rozin’s insight is that the pleasure of capsaicin is not merely the endorphin afterglow that follows the pain; it is the simultaneous awareness that the pain is real and the danger is not. The higher-order cortex—the evaluative, contextualizing brain—monitors the TRPV1 alarm and performs a rapid override: the mouth is screaming “fire,” but there is no fire; the nociceptive system is reporting tissue damage, but no tissue is being damaged. It is this cognitive gap between the body’s conviction and the mind’s knowledge that generates the characteristic thrill. The pleasure derives, in significant part, from the psychological satisfaction of having tricked one’s own evolutionary hardware—a felt sense of mastery over the body’s most ancient and authoritative warning system. The eater is not merely enduring pain; they are staging a private rebellion against the tyranny of their own reflexes, and winning.
What Rozin identified, in effect, is the structure of a “safe threat”—an experience that provides all the physiological markers of a genuine biological crisis (the racing heart, the sweating, the flood of stress hormones) without any of the biological cost. This is the same architecture that underlies the appeal of roller coasters, horror films, and skydiving: the body is fully mobilized for an emergency that the mind knows will not arrive. But capsaicin occupies a unique position within this category, because the threat is not external and environmental but internal and somatic—the danger signal originates from inside the body itself, from the very tissue of the mouth, making the override feel more intimate, more like a genuine contest between self and self. This psychological dimension is the critical bridge between the biological story told above and the social story that follows. The endorphin loop explains how pain becomes tolerable; benign masochism explains how it becomes desirable. And it is precisely at the moment when the individual discovers this private pleasure—this quiet triumph of cognition over nociception, of the knowing mind over the screaming body—that the experience becomes available for social interpretation. The internal mastery must be performed to be witnessed, and once it is witnessed, it enters the semiotic field. The private rebellion becomes a public signal. The body’s honest alarm, already overridden by the individual’s own cognition, is now ready to be overwritten again—this time by the collective scripts of culture, status, and identity.
The Social Layer and the Semiotic Field
This is where the “Social Poison” of interpretive overfitting begins. In a social context, we rarely allow a physiological reaction to simply be; we over-interpret it, assigning intent and character to involuntary biological processes. The raw data of a sweating brow is overwritten by cultural scripts, leading to two primary, often conflicting, narratives.
The Performance Hypothesis
The first narrative is the Performance Hypothesis, which views the consumption of extreme heat as a calculated display of status or belonging. In this framework, the visible struggle with spice is seen as a performative act—a “masochistic” display designed to signal toughness, worldliness, or membership in a specific subculture (such as the “chili-head” community). Here, the “honesty” of the biological response is ironically subverted; the very fact that the reaction is involuntary makes it the perfect stage for a performance of endurance. The consumer isn’t just eating; they are demonstrating their ability to colonize their own biological pain for social capital.
What gives this performance its peculiar credibility is a dynamic that biologists call the Handicap Principle and game theorists call costly signaling: a signal is trustworthy precisely because it is expensive to produce. A peacock’s tail is convincing evidence of genetic fitness because growing and carrying it is a genuine metabolic burden; a bluff would collapse under the weight. The spicy-food performance operates on the same logic. The visible struggle—the flushed skin, the watering eyes, the involuntary gasp—is not a flaw in the display; it is the display. It works as a status signal precisely because it hurts. The pain is the price of admission, and only those who value the resulting social capital enough to pay it, or who possess a naturally high capsaicin tolerance that lowers the cost, will sit down at the table and order the Carolina Reaper. Someone who merely claims to love extreme heat but blanches at the first bite gains nothing; the body’s involuntary betrayal exposes the bluff instantly. This is the deep irony at the heart of the Performance Hypothesis: the biological honesty of the capsaicin response—the very thing that seems to undercut a polished social performance—is what makes the performance credible in the first place. Because the cost cannot be faked, the signal cannot be counterfeited. The sweating brow is not a failure of composure; it is the receipt that proves the toll was paid.
The Toughness Hypothesis
The second narrative is the Toughness Hypothesis, which interprets spice tolerance as a proxy for moral or physical character. In many cultures, the ability to “handle” heat is conflated with grit, resilience, and even masculinity. Under this lens, the person who does not sweat or flush is seen as possessing a superior constitution. The biological variation in TRPV1 receptor density—a matter of genetic luck—is transformed into a moral trait. To have a high tolerance is to be “strong”; to struggle is to be “weak.” This hypothesis ignores the biological reality of desensitization and genetics, replacing it with a semiotic system where the tongue becomes a site of character judgment.
From a psychological standpoint, this conflation of tolerance with character is a textbook instance of the Fundamental Attribution Error—the well-documented tendency to attribute others’ behavior to stable dispositional traits (courage, weakness, discipline) while underweighting the situational and biological variables that actually explain the behavior. When an observer watches someone eat a scorpion pepper without flinching and concludes that this person is tough, they are performing precisely the attribution error that Lee Ross identified in 1977: mistaking a situational outcome—one shaped by prior exposure history, cultural habituation, and, crucially, genetic endowment—for a window into the person’s character. The error is compounded by the fact that the biological substrate in question is not uniform across the population. The TRPV1 gene exhibits significant polymorphic variation: single-nucleotide polymorphisms (SNPs) in and around the TRPV1 locus alter receptor expression levels, ion-channel sensitivity, and downstream signal amplification. Some individuals are born with a sparse distribution of functional TRPV1 receptors in their oral mucosa; others carry variants that produce receptors with a higher activation threshold, requiring more capsaicin to trigger the same nociceptive cascade. What society codes as “strength”—the calm, unflushed consumption of extreme heat—may therefore be, in the most literal sense, a partial absence of sensory hardware: not a triumph of will over pain, but a quieter peripheral signal that never reached the volume the observer imagines was heroically suppressed. Conversely, the person who gasps and reaches for milk may possess a denser receptor field or a more sensitive channel variant—a richer sensory apparatus, not a weaker constitution. The Toughness Hypothesis thus performs a double inversion: it reads a genetic variable as a moral constant, and it rewards sensory deficit as though it were psychological surplus. By mapping the Fundamental Attribution Error onto the biology of TRPV1 polymorphism, we can see that the entire edifice of spice-as-character-test rests on a failure to distinguish between what the body is and what the person chooses—between the hand one is dealt and the way one plays it.
The Sensory Semiotic Triad: Spicy, Sour, and Bitter
To fully appreciate the unique position of heat in the social hierarchy, we must compare it to other intense sensory experiences. We can conceptualize a Sensory Semiotic Triad consisting of the spicy, the sour, and the bitter. While all three involve a degree of “acquired” appreciation, they occupy vastly different roles in the theater of social interaction.
Spicy and sour sensations are inherently public dramas. Both trigger immediate, involuntary, and highly visible physiological responses—the “sour face” (puckering of the lips, squinting of the eyes) and the “spicy flush” (sweating, gasping, redness). These signals are ambiguous; they can represent either distress or intense pleasure, creating a semiotic gap that the observer must fill. Crucially, these signals are what semioticians in the Peircean tradition would call polysemic—they carry multiple, competing meanings simultaneously. A flushed face and watering eyes could signify genuine agony, masochistic pleasure, performative bravado, or the threshold moment between surrender and triumph. The observer cannot determine which meaning is “correct” from the signal alone, and it is precisely this semiotic ambiguity that serves as the engine of social co-optation. Because the sign underdetermines its meaning, it invites interpretation—and where interpretation is invited, social narratives rush in. The “sour challenge” or the “hot wing challenge” relies on this polysemic visibility to turn a private sensation into a shared spectacle, a text that the audience actively co-authors.
In contrast, bitterness remains largely a private signal. While a profound bitterness might cause a brief grimace, it lacks the sustained, escalating physical drama of heat or the sharp, contorting reflex of sourness—there is no progressive arc of visible suffering, no crescendo for an audience to track. More fundamentally, the bitter response is monosemic: it means one thing—rejection. The grimace at a bitter compound is read, almost universally, as aversion. There is no interpretive gap, no ambiguity about whether the person is suffering beautifully or performing resilience. The sign arrives pre-interpreted, and where there is no interpretive gap, there is no space for social narrative to colonize. A polysemic signal is an open text; a monosemic signal is a closed one. No one watches someone sip an intensely bitter espresso and wonders, “Is she in pain or ecstasy? Is this a feat of endurance?”—the grammar of the response simply does not pose the question.
Bitterness is therefore processed as a matter of internal preference—a “quiet” taste that one either enjoys or avoids, a private dialogue between the palate and the brain rather than a public broadcast of the body’s resilience. There is little social capital to be gained from “enduring” a bitter radicchio or a dark coffee in the same way one “conquers” a ghost pepper, because the bodily response to bitterness is neither legible enough to be read at a distance nor ambiguous enough to sustain competing interpretations. It is a preference, not a feat. The Triad thus reveals a principle: the social life of a sensation is proportional to the polysemy of its signal. Spicy and sour sensations become theaters of identity precisely because their signs are open to multiple readings; bitterness remains a private affair precisely because its sign is closed. The engine of social co-optation is not intensity alone—it is interpretive possibility.
Cultural Metastasis and Status Games
As the individual performance of spice consumption scales, it metastasizes into broader cultural tropes and identity politics. The semiotic field expands from the personal to the collective, where heat becomes a form of cultural capital. In this arena, the ability to navigate extreme spice is no longer just about personal toughness; it is a signal of worldliness and “authentic” engagement with the Other.
Spice as Cultural Capital and Authenticity Policing
In the globalized culinary market, spice levels are often used as a metric for authenticity. This leads to a form of authenticity policing, where the “true” version of a dish is defined by its proximity to a perceived threshold of pain. Diners may demand “Thai spicy” or “Indian spicy” as a way to distance themselves from the “diluted” or “Westernized” versions of a cuisine. Here, the heat is not just a flavor profile but a gatekeeping mechanism. To consume the heat is to prove one’s worthiness to participate in the culture, while the inability to do so is dismissed as a lack of culinary sophistication or a sheltered upbringing.
Yet this gatekeeping mechanism carries a deeper distortion that culinary scholars have identified as culinary orientalism: the reduction of an entire cuisine’s vast internal diversity to a single painful metric. To demand “Thai spicy” is to flatten a tradition in which many canonical dishes—the coconut-milk richness of tom kha gai, the delicate balance of pad see ew, the intentional mildness of a kanom jeen with mild fish curry—are designed to be gentle precisely because they provide contrast and balance within a meal structured around complementary sensations. Thai cuisine, like most complex culinary traditions, is an architecture of counterpoints: hot against cool, rich against sharp, intense against mild. To define its “authenticity” by its maximum capsaicin output is to mistake one voice in a choir for the entire performance—and, in doing so, to erase the compositional intelligence that makes the tradition worth celebrating in the first place. The semiotic field does not merely distort social dynamics; it flattens the very cuisines it claims to honor.
The narrowing runs deeper still. The semiotic field’s fixation on capsaicin-based heat as the metric of spice courage ignores the fact that pungency itself is a diverse sensory landscape. The numbing, electric mā (麻) of Sichuan peppercorns—produced by hydroxy-alpha-sanshool acting on mechanoreceptors rather than thermoreceptors—offers a form of intensity that is neurologically distinct from the capsaicin burn: it tingles, buzzes, and temporarily rewires the mouth’s sense of touch rather than its sense of temperature. The sharp nasal pungency of wasabi and horseradish, mediated by allyl isothiocyanate activating TRPA1 receptors in the sinuses, produces a fleeting, almost cerebral flash of pain that dissipates in seconds—a sensation with no sustained arc of visible suffering and therefore little utility for the endurance performances the semiotic field rewards. These alternative pungencies offer extraordinary complexity and cultural depth, yet they remain marginal to the dominant spice discourse precisely because they do not produce the prolonged, legible, polysemic spectacle that capsaicin does. The semiotic field, in other words, has not merely co-opted heat as a social signal; it has selected for a specific kind of heat—the kind that best serves performative endurance—and in doing so, has narrowed the sensory landscape even as it claims to celebrate adventurousness.
The Sophistication Paradox
This leads to the sophistication paradox: the claim that a high tolerance for heat signals a more refined and adventurous palate. Proponents of this view argue that spice enhances the complexity of a dish, yet at extreme levels, capsaicin objectively obliterates flavor nuance by overwhelming the taste buds and triggering a pain response that drowns out subtle aromatics. The paradox lies in the fact that the “refined” palate is often celebrating the very thing that prevents it from tasting the food’s finer details. The heat becomes a proxy for sophistication, even as it acts as a sensory blackout curtain—and the claim that extreme spice enhances one’s appreciation of a dish is, as a culinary analogy makes vivid, like saying you appreciate music more because you listen to it at 120 decibels. At that volume, the subtlety of the composition is not revealed but annihilated; what remains is not discernment but endurance. Game theory sharpens the absurdity further. Within the spicy food semiotic field, social capital is often valued more than the primary utility of the food itself, and the sophistication paradox is the point where this priority structure becomes fully visible. The consumer who orders the hottest item on the menu is executing a strategic trade-off: sacrificing the utility of flavor—the very thing that ostensibly defines culinary sophistication—in order to gain the utility of status. This is not a hidden cost; it is the entire mechanism. The diner pays in destroyed nuance and collects in social capital. Yet the sophistication claim requires that this trade-off remain unacknowledged, because the moment it is made explicit, the paradox collapses into plain contradiction: one cannot simultaneously claim to be maximizing gustatory appreciation while pursuing a strategy that systematically minimizes it. The “refined palate” is, in game-theoretic terms, a player who has optimized for a payoff (status) that is structurally incompatible with the payoff they advertise (taste)—and the sensory blackout curtain is not an unfortunate side effect of their strategy but its operative mechanism.
The Mayonnaise Proxy
Conversely, the absence of spice is weaponized through the “mayonnaise” stereotype. In contemporary social discourse, “mayonnaise” and “blandness” have become linguistic proxies for a specific type of identity—often associated with whiteness, lack of culture, or a fragile, unadventurous constitution. This stereotype functions as the inverse of the toughness hypothesis; if heat is grit, then mildness is weakness. By reducing complex cultural identities to their perceived tolerance for capsaicin, the semiotic field transforms a condiment into a tool for social categorization and identity-based derision. The “mayonnaise” label suggests a sensory sterility that is equated with a lack of historical or cultural depth, further entrenching the spicy food semiotic field as a primary site of modern social signaling.
The Industrialization of Suffering
The final stage in the evolution of the spicy food semiotic field is its commercialization and media-driven escalation—what might be termed the Hot-Wing Industrial Complex. In this phase, the private biological struggle is commodified and transformed into a public spectacle. Media platforms, most notably YouTube series like Hot Ones, have pioneered a new genre of interview where the physiological distress of the subject is the primary draw. The “biological honesty” of the capsaicin response is leveraged as a tool for “authentic” celebrity exposure; the theory being that a person cannot maintain a PR-managed facade while their TRPV1 receptors are screaming.
This industrialization relies heavily on Scoville-based marketing, where the heat level of a product is no longer a culinary choice but a competitive metric. Sauces are marketed with names that evoke violence, death, or insanity, turning the grocery aisle into a gauntlet of performative bravery. This turns the act of eating into a “challenge”—a discrete event designed for social media documentation. The biological struggle is no longer a byproduct of a meal; it is the product itself. Game theory reveals the structural logic driving this escalation. The Mayonnaise Proxy penalty described above—the social cost of being seen to prefer mild food—functions as a powerful negative payoff that reshapes the entire decision landscape. When the stigma attached to choosing mildness is sufficiently severe, the semiotic field collapses into what game theorists call a Pooling Equilibrium: a state in which all players, regardless of their private type (their actual capsaicin tolerance or preference), choose the same publicly visible action—extreme heat—because the cost of revealing oneself as mild-preferring exceeds the physical cost of enduring the burn. High-tolerance and low-tolerance individuals alike converge on the same fiery order, not because they share a preference but because the signaling environment has made the alternative socially untenable. The menu ceases to be a space of genuine choice and becomes a one-dimensional race to the top of the Scoville scale, where the only “safe” move is to match or exceed the group’s current threshold.
This pooling dynamic generates a Prisoner’s Dilemma at the heart of every shared meal. Consider two diners at a table: each might privately prefer a milder dish—one that would allow them to taste the food’s actual flavor profile and avoid the gastrointestinal aftermath. Yet each knows that choosing mildness while the other chooses heat will trigger the Mayonnaise Proxy penalty, coding them as the “weak” one in the pair. The dominant strategy for both players, therefore, is to choose spicy—not because it maximizes their individual gustatory utility, but because it minimizes their social risk. The result is a Pareto-inferior equilibrium: both diners end up suffering more than necessary, consuming food that overwhelms their palates and punishes their digestive tracts, when a world in which both chose mildly would have left each of them better off on every dimension except the one that matters in the semiotic field—status. The tragedy is structural, not personal. No single diner can unilaterally defect to mildness without bearing the full weight of the social penalty, and so the equilibrium holds, meal after meal, even though every participant in it would prefer a different world.
The commercial arm of the Hot-Wing Industrial Complex is locked into a parallel escalation that evolutionary biologists would recognize as a Red Queen race—the dynamic in which organisms must constantly evolve merely to maintain their current fitness relative to co-evolving competitors. As the pooling equilibrium drives consumers toward ever-higher Scoville counts, and as repeated exposure triggers the chili bleaching and TRPV1 desensitization described earlier, the spectacle’s impact erodes. A sauce that once produced dramatic, camera-ready suffering becomes, through habituation, merely uncomfortable—and a merely uncomfortable sauce generates no social media content, no viral moments, no signaling value. Brands must therefore innovate relentlessly toward more extreme formulations—higher capsaicinoid concentrations, novel cultivars like Pepper X and Apollo, synthetic capsaicin analogs—not to increase the level of social signaling their products provide, but simply to maintain it against the rising baseline of consumer tolerance. They are running to stand still. The result is an arms race between the human nervous system’s capacity for adaptation and the food industry’s capacity for chemical escalation, each driving the other forward in a feedback loop that has no natural resting point. The Scoville number on the label is not a measure of flavor; it is an index of how far the race has run.
Furthermore, this spectacle creates a new form of social cohesion: the ritual of suffering together. Whether in a viral “One Chip Challenge” or a group outing to a wing joint with a “wall of flame,” the shared experience of intense, non-lethal pain acts as a powerful bonding mechanism. This is a secular, sensory ritual where the participants prove their mettle and their shared identity through a collective descent into physiological chaos. The pooling equilibrium ensures that the ritual’s entry price keeps rising—what counted as a credible performance of toughness five years ago is now the table stakes, and the Prisoner’s Dilemma ensures that no individual participant can opt for a gentler experience without defecting from the group’s implicit social contract. By turning the burn into a stage for performing identity, and by locking both consumers and producers into interlocking escalation dynamics—the former trapped in a Pareto-inferior social equilibrium, the latter sprinting on the Red Queen’s treadmill—the Hot-Wing Industrial Complex ensures that the capsaicin paradox remains a central, if increasingly manufactured and increasingly painful, pillar of modern social interaction.
Conclusion: The Body as a Semiotic Surface
The journey from the TRPV1 receptor to the “Hot-Wing Industrial Complex” illustrates that the human body is never a closed biological system. It is a semiotic surface—a canvas upon which society projects its values of toughness, authenticity, and status. The “biological honesty” of a flush or a tear is immediately captured and recoded by the “social poison” of interpretation.
What this entire apparatus amounts to is a domestication of the absolute. The capsaicin response begins as a biological absolute—a raw, unmediated chemical scream from the TRPV1 receptor, a signal that evolved to mean one thing and one thing only: danger, stop, do not eat this. Yet we take this absolute and domesticate it, breaking it like a wild animal to the harness of social utility. We train it to mean toughness, authenticity, worldliness, belonging. We build elaborate semiotic architectures—the Performance Hypothesis, the Toughness Hypothesis, the Sophistication Paradox, the Hot-Wing Industrial Complex—all of which are, at bottom, cultural infrastructure for converting a biological emergency signal into a legible social currency. The “honesty” we attribute to the body’s involuntary response is not a property we discover in nature; it is a cultural layer we apply, a domesticating frame that makes sense of our peculiar desire to suffer for pleasure, for status, for the feeling of being fully alive in a body we can narrate to others. We do not honor the biological signal by reading it as honest; we tame it by doing so, rendering it safe enough to circulate in the economy of social meaning.
But domestication has a boundary, and that boundary reveals something essential about the entire enterprise. There exists a threshold—what we might call the point of semiotic collapse—where pain becomes so overwhelming that the social field dissolves entirely. Elaine Scarry, in The Body in Pain (1985), argued that intense physical pain is fundamentally world-destroying: it annihilates language, unmakes the sufferer’s capacity for symbolic thought, and reduces the human subject to pre-linguistic cries that carry no social meaning whatsoever. At the extreme end of the capsaicin spectrum—the point where a Carolina Reaper or a concentrated extract overwhelms not just the palate but the entire nervous system—we approach this threshold. The sufferer no longer performs toughness or signals authenticity; they simply hurt, in a way that is illegible to any semiotic framework. The flush is no longer a polysemic sign open to competing interpretations; it is the body in revolt, beyond narrative, beyond performance, beyond meaning. Language fails. The groan that escapes is not a social signal but a biological reversion—the organism stripped of its cultural clothing, returned to the pre-symbolic animal cry that no audience can decode as anything other than what it is.
The entire spicy food semiotic field, then, operates in the narrow space between two poles: the embodiment anchor—the point where capsaicin forces consciousness back into the body and makes the flesh available for social narration—and the threshold of semiotic collapse—the point where pain overwhelms all narration and the body ceases to be a legible text. Every hot wing challenge, every “Thai spicy” request, every Scoville-escalation arms race is a controlled flirtation with the biological real: an approach toward the abyss of unsymbolizable pain that never quite tips over the edge. The social utility of the burn depends entirely on maintaining this balance—enough pain to produce the honest, costly signal that the semiotic field requires, but not so much that the signal dissolves into the formless scream that no culture can metabolize. The domestication of the absolute is, in this light, an act of extraordinary precision: we must bring the wild animal close enough to feel its heat, but not so close that it devours us.
This machinery is not unique to capsaicin. We see identical patterns in the rise of extreme wellness rituals, such as the “cold plunge” or high-intensity interval training. In these contexts, the physiological shock—the “cold shock response”—is marketed and performed as a sign of superior discipline and mental fortitude. The shivering body, much like the sweating tongue, becomes a site of moral signaling. The raw data of the nervous system is consistently overwritten by the software of social expectation. And in each case, the same two boundaries apply: the experience must be intense enough to anchor the body as a legible semiotic surface, but not so intense that it crosses into the territory Scarry mapped—the zone where pain unmakes the world and the social field goes dark.
Ultimately, the capsaicin paradox reveals a fundamental human drive: the need to metabolize raw, chaotic sensation into structured, social meaning. We do not merely inhabit our bodies; we narrate them. But the narration is always precarious, always conducted at the edge of a silence that would swallow it whole. By transforming a plant’s defense mechanism into a cultural metric, we demonstrate that even our most visceral pains are never truly our own—they are always already part of the stories we tell about who we are and what we value. Yet the stories hold only so long as the pain remains domesticated. Beyond the last Scoville unit that language can metabolize lies the biological real—mute, absolute, and indifferent to every meaning we have tried to make it carry.
Comic Book Generation Task
Generated Script
Full Script
The Capsaicin Paradox
A philosophical and physiological descent into the “Spicy Food Semiotic Field,” exploring how a plant’s defense mechanism became a tool for human status, performance, and the eventual annihilation of language itself.
Characters
- Dr. Aris Thorne: Cynical, intellectual, detached observer of human folly. Narrator/Philosopher. The guide for the reader through the ‘Semiotic Field’. (A sharp-featured academic in a dark turtleneck and a lab coat that looks like a trench coat. Often seen in silhouette or half-shadow, holding a single chili pepper like a skull in a Hamlet soliloquy.)
- The Subject (Alex): Initially seeking social capital and ‘authenticity’, eventually reduced to primal biological reaction. Protagonist/Victim. The test subject for Dr. Thorne’s theories. (An average person whose appearance shifts from composed and trendy to a distorted, sweating, red-faced mess as the comic progresses. Wears a ‘Spice Seeker’ t-shirt.)
Script
Page 1
Row 1
- Panel 1: Close up on Dr. Thorne’s face, half-hidden in shadow. He holds a bright red Habanero between his thumb and forefinger.
- Dr. Thorne: “Meet the Capsicum. It doesn’t want to be eaten by you.”
- Caption: It begins with a lie. A molecular deception.
- Panel 2: A technical overlay of the Capsaicin molecule glowing in neon orange.
- Caption: It engineered a key. A molecular shape designed to fit a very specific lock. Row 2
- Panel 1: A surreal diagram of a human mouth. The TRPV1 receptors are depicted as glowing “fire alarms” on the tongue.
- Dr. Thorne: “The TRPV1 receptor. A thermosensor calibrated for 43°C. It’s there to tell you your mouth is on fire.”
- Panel 2: The Capsaicin molecule “plugging into” the receptor. Sparks of neon red fly off.
- Dr. Thorne: “The molecule lies. It tells the brain you are burning, even when the temperature hasn’t moved a degree.”
- Caption: DIRECTED DETERRENCE. Row 3
- Panel 1: A monochrome bird eating a chili pepper, looking completely unbothered.
- Dr. Thorne: “Birds lack the lock. They eat, they fly, they disperse the seeds. The plant’s reproductive future is secure.”
- Panel 2: Dr. Thorne looks directly at the reader, the chili pepper now closer to his own mouth.
- Dr. Thorne: “But humans? We heard the alarm… and we decided we liked the sound.”
Page 2
Row 1
- Dr. Thorne: “But humans? We heard the alarm… and we decided we liked the sound.”
- Panel 1: Alex (The Subject) sits at a lab table. He takes a bite of a pepper. He looks confident.
- Alex: “I can handle it. I’m a “Spice Seeker.””
- Panel 2: Close up on Alex’s eye. The pupil dilates. A single bead of sweat forms.
- Caption: BIOLOGICAL HONESTY. Row 2
- Panel 1: A split-screen. On one side, Alex’s brain is a chaotic mess of “FIRE!” warnings. On the other, a calm Dr. Thorne stands in a library.
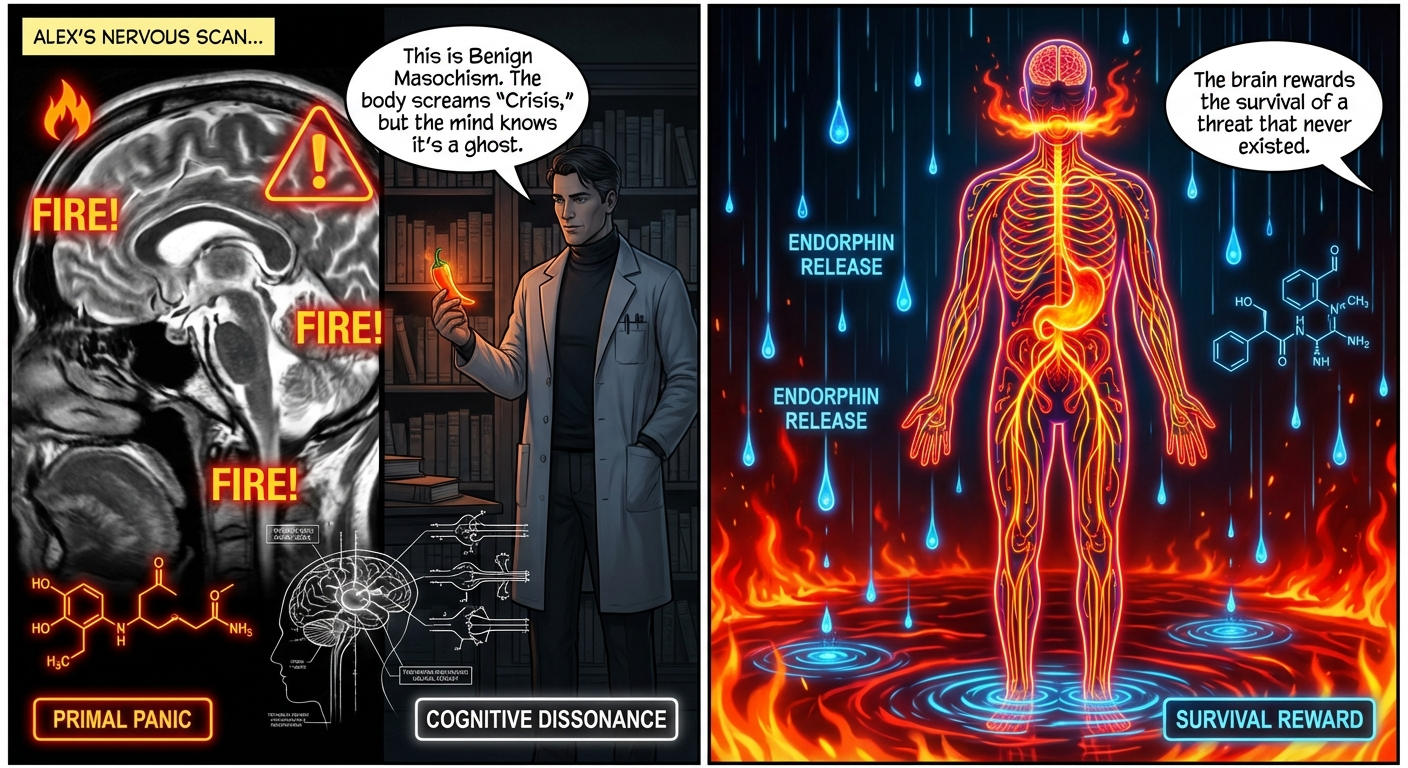
- Dr. Thorne: “This is Benign Masochism. The body screams “Crisis,” but the mind knows it’s a ghost.”
- Panel 2: Alex’s nervous system is lit up like a neon circuit board. Endorphins are depicted as glowing blue droplets falling into a sea of red fire.
- Dr. Thorne: “The brain rewards the survival of a threat that never existed.”
- Caption: THE ENDORPHIN LOOP. Row 3
- Panel 1: Alex is now smiling through the pain, though his face is turning a light neon pink.
- Alex: “(Thinking) This… this feels amazing.”
- Panel 2: Dr. Thorne leans over Alex, looking like a shadow.
- Dr. Thorne: “You aren’t just eating, Alex. You’re staging a rebellion against your own evolutionary hardware. And you think you’re winning.”
Page 3
Row 1
- Dr. Thorne: “You aren’t just eating, Alex. You’re staging a rebellion against your own evolutionary hardware. And you think you’re winning.”
- Panel 1: Alex is at a table with friends. He’s eating a “Nuclear Wing.” He is visibly struggling—sweating, red-faced—but he’s holding a thumb up.
- Caption: THE PERFORMANCE HYPOTHESIS.
- Panel 2: Dr. Thorne stands in the background, leaning against a pillar, watching.
- Dr. Thorne: “The pain is the price of admission. If it didn’t hurt, it wouldn’t be a signal.” Row 2
- Panel 1: A diagram of a peacock’s tail overlays the restaurant scene.
- Dr. Thorne: “Biologists call it the Handicap Principle. A signal is only honest if it’s expensive.”
- Panel 2: Close up on Alex’s sweating brow. The sweat drops are labeled “THE RECEIPT.”
- Dr. Thorne: “You can’t fake the flush, Alex. That’s why they believe you’re “tough.”” Row 3
- Panel 1: Alex’s friends are cheering, but their faces are slightly distorted, like masks.
- Friend: “Man, you’re a beast! I could never do that.”
- Panel 2: Dr. Thorne holds a magnifying glass up to the scene.
- Dr. Thorne: “The biological event is over. Now, it’s just Social Poison. Interpretation has begun.”
Page 4
Row 1
- Dr. Thorne: “The biological event is over. Now, it’s just Social Poison. Interpretation has begun.”
- Panel 1: A comparison of two tongues. One has many glowing TRPV1 receptors (labeled “SENSITIVE”), the other has very few (labeled “BLEACHED”).
- Dr. Thorne: “Some are born with a sparse field. Others have burned their receptors into silence.”
- Panel 2: Alex is looking at a “Mayonnaise” jar with a look of disgust.
- Caption: THE MAYONNAISE PROXY. Row 2
- Panel 1: Alex is looking down at a person eating a mild korma. Alex looks “superior.”
- Alex: “Some people just don’t have the grit for real food.”
- Panel 2: Dr. Thorne points to a DNA helix.
- Dr. Thorne: “You’re mistaking a sensory deficit for a moral surplus, Alex. It’s not character; it’s polymorphism.” Row 3
- Panel 1: Alex is eating something so hot it’s literally glowing. He can’t see the food anymore; it’s just a white light.
- Caption: THE SENSORY BLACKOUT CURTAIN.
- Panel 2: Dr. Thorne sips a glass of water, looking bored.
- Dr. Thorne: “You claim it “enhances” the flavor. But you’re listening to Mozart at 120 decibels. You aren’t hearing the music; you’re just enduring the noise.”
Page 5
Row 1
- Dr. Thorne: “You claim it “enhances” the flavor. But you’re listening to Mozart at 120 decibels. You aren’t hearing the music; you’re just enduring the noise.”
- Panel 1: Alex is now a guest on a YouTube show. Cameras are everywhere. The host is a silhouette.
- Host: “Welcome to the Gauntlet! Ready for the “Reaper’s Breath”?”
- Panel 2: A wall of hot sauce bottles with names like “SUDDEN DEATH” and “COLON COLLAPSE.”
- Caption: SCOVILLE-BASED MARKETING. Row 2
- Panel 1: A graph showing Scoville levels skyrocketing over the years.
- Dr. Thorne: “To keep the signal “honest,” the heat must rise. You’re running to stand still.”
- Panel 2: Alex takes a bite. His face begins to melt into a smear of neon orange.
- Caption: THE POOLING EQUILIBRIUM. Row 3
- Panel 1: Two people at a table, both looking terrified of the wings in front of them, but neither will back down.
- Diner 1: “(Thinking) I hate this.”
- Diner 2: “(Thinking) I’m going to die.”
- Both: ““This is great, right?””
- Panel 2: Dr. Thorne stands between them, arms crossed.
- Dr. Thorne: “A Pareto-inferior equilibrium. You’re both suffering because you’re afraid of a condiment.”
Page 6
Row 1
- Dr. Thorne: “A Pareto-inferior equilibrium. You’re both suffering because you’re afraid of a condiment.”
- Panel 1: Alex is clutching the table. His eyes are streaming. He tries to speak, but only a jagged, neon-red shape comes out of his mouth.
- Alex: “I… I… [UNINTELLIGIBLE]”
- Panel 2: Dr. Thorne stands in a void of pure white.
- Dr. Thorne: “We’ve reached the point of Semiotic Collapse. The world is being unmade.” Row 2
- Panel 1: The restaurant, the cameras, the friends—all are dissolving into a sea of fire. Only Alex’s agonized face remains.
- Caption: PAIN ANNIHILATES LANGUAGE.
- Panel 2: Close up on Alex’s mouth. The word “AUTHENTIC” is shattering into pieces.
- Dr. Thorne: “The “Biological Real” has returned. It doesn’t care about your status or your “Thai Spicy” request.” Row 3
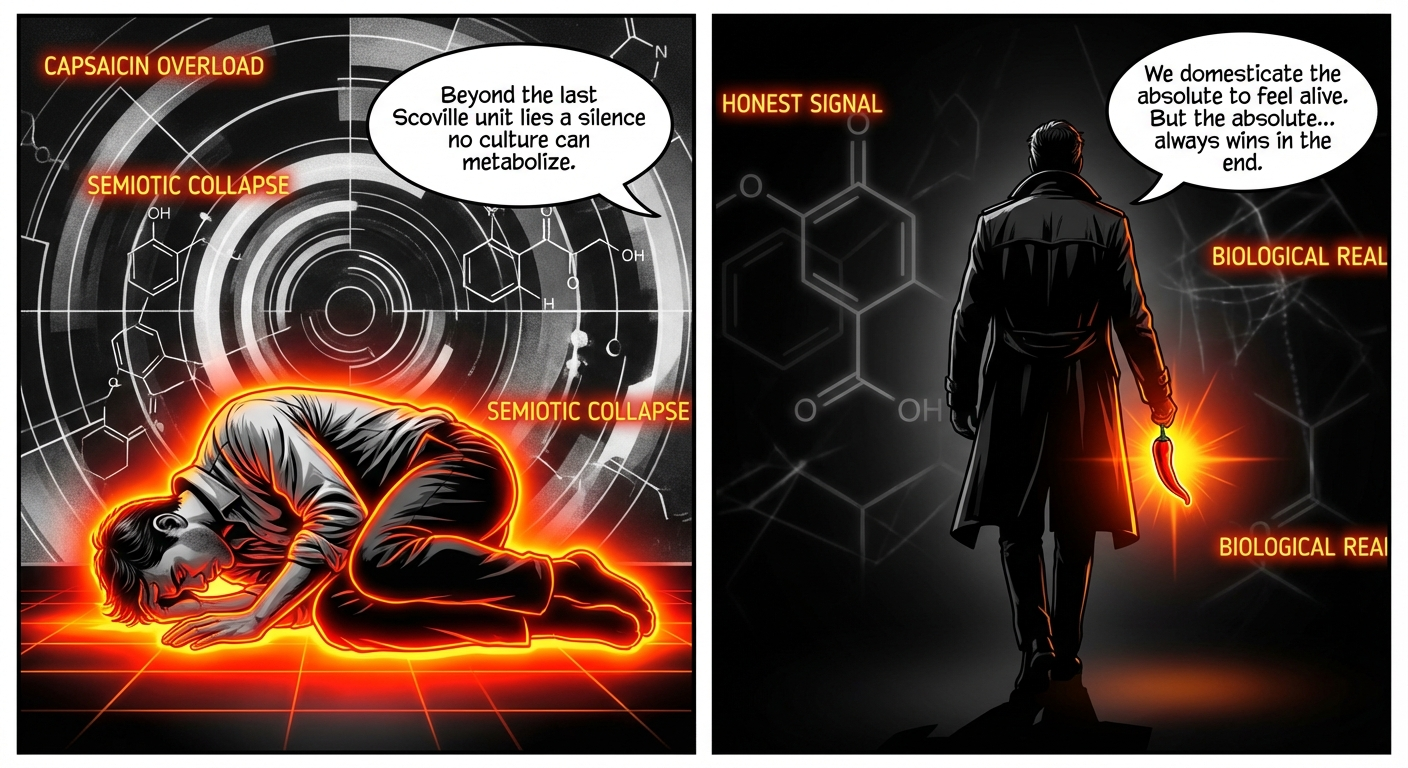
- Panel 1: Alex is curled in a fetal position on a floor of pure neon orange. He is silent.
- Dr. Thorne: “Beyond the last Scoville unit lies a silence no culture can metabolize.”
- Panel 2: Dr. Thorne walks away into the darkness, leaving the reader with a final look at the red chili pepper, now glowing like a dying star.
- Dr. Thorne: “We domesticate the absolute to feel alive. But the absolute… always wins in the end.”
- Caption: (Small, fading text) The body is a surface. The burn is the truth.
Auto-Fix Mode
Auto-applying: proceeding directly to visual generation.
Character: Dr. Aris Thorne

Cynical, intellectual, detached observer of human folly. Narrator/Philosopher. The guide for the reader through the ‘Semiotic Field’.
Character: The Subject (Alex)

Initially seeking social capital and ‘authenticity’, eventually reduced to primal biological reaction. Protagonist/Victim. The test subject for Dr. Thorne’s theories.
















Narrative Generation Task
Overview
Narrative Generation
Subject: The Scoville Gauntlet: A Dramatization of the Spicy Food Semiotic Field
Configuration
- Target Word Count: 2500
- Structure: 3 acts, ~2 scenes per act
- Writing Style: literary
- Point of View: third person limited
- Tone: dramatic
- Detailed Descriptions: ✓
- Include Dialogue: ✓
- Internal Thoughts: ✓
Started: 2026-02-20 20:56:57
Progress
Phase 1: Narrative Analysis
Running base narrative reasoning analysis…
Cover Image
Prompt:

High-Level Outline
The Scoville Gauntlet
Premise: An exploration of the intersection of biological reality and social artifice through the lens of extreme capsaicin consumption, where a cultural critic and a lifestyle influencer battle to maintain their public personas under intense physical duress.
Estimated Word Count: 2496
Characters
Elias Thorne
Role: protagonist
Description: A lean, forty-something cultural critic with salt-and-pepper hair and a wardrobe of expensive, neutral-toned linens. He possesses a ‘curated’ face—one practiced in the art of the unimpressed smirk.
Traits: Intellectual, prideful, and deeply terrified of being seen as ‘common.’ He views the Gauntlet as a semiotic challenge—a test of his ability to maintain a sophisticated persona while his nervous system is under siege. He wants to prove that the mind can colonize the body’s pain.
Maya Vance
Role: antagonist / foil
Description: A twenty-four-year-old ‘stunt-lifestyle’ influencer. She is vibrant, wearing neon athletic gear that matches the studio lights. Her movements are twitchy, energetic, and calculated for maximum engagement.
Traits: Pragmatic, competitive, and hyper-aware of the ‘economy of attention.’ She views pain as a currency. Her goal is not to hide the suffering, but to weaponize it—to perform ‘authenticity’ more effectively than Elias performs ‘sophistication.’
Marcus “The Architect” Reed
Role: supporting / catalyst
Description: The host of the show. He wears a clinical white lab coat over a black turtleneck. He speaks in a soothing, low-frequency baritone that contrasts with the violence of the food.
Traits: Detached, observant, and slightly sadistic. He represents the ‘Industrialized Suffering’ of the media machine. He doesn’t care who wins; he only cares about the ‘spicy flush’ captured in 8K resolution.
Settings
gauntlet_studio
Description: A soundstage dominated by deep obsidian blacks and piercing neon magentas. The center table is a slab of polished white quartz, reflecting the high-intensity LED arrays overhead. Cameras are positioned at ‘predatory’ angles—extreme close-ups designed to capture sweat, pupil dilation, and the micro-tremors of the lips.
Atmosphere: Clinical, claustrophobic, and predatory. It feels less like a kitchen and more like a high-tech interrogation room or a biological laboratory.
Significance: It represents the ‘Panopticon of Performance,’ where every biological betrayal is recorded and commodified.
Act Structure
Act 1: The Illusion of Control
Purpose: To establish the ‘Sophistication Paradox’ and the initial social posturing between Elias and Maya.
Estimated Scenes: 2
Key Developments:
- The introduction of the first two wings
- Elias’s attempt to intellectualize the flavor profiles
- The first signs of physiological stress
Act 2: The Semiotic Collapse
Purpose: To dramatize the ‘Prisoner’s Dilemma’ of pain and the breakdown of the characters’ public personas.
Estimated Scenes: 2
Key Developments:
- The transition into the ‘danger zone’ of Scoville units
- The loss of verbal eloquence
- The physical ‘Red Queen Race’ where neither can afford to stop
Act 3: The Altar of the Scoville
Purpose: To reach the ‘Threshold of Semiotic Collapse’ and resolve the conflict through a moment of ‘Biological Honesty.’
Estimated Scenes: 2
Key Developments:
- The final wing consumption
- The climax of the physical pain
- The aftermath where the cameras stop rolling and personas dissolve
Status: ✅ Pass 1 Complete
Outline
The Scoville Gauntlet
Premise: An exploration of the intersection of biological reality and social artifice through the lens of extreme capsaicin consumption, where a cultural critic and a lifestyle influencer battle to maintain their public personas under intense physical duress.
Estimated Word Count: 2496
Total Scenes: 6
Detailed Scene Breakdown
Act 1: The Illusion of Control
Purpose: To establish the ‘Sophistication Paradox’ and the initial social posturing between Elias and Maya. Key Developments: The introduction of the first two wings; Elias’s attempt to intellectualize the flavor profiles; the first signs of physiological stress.
Scene 1: The Semiotic Appetizer
- Setting: gauntlet_studio
- Characters: Elias Thorne, Maya Vance, Marcus “The Architect” Reed
- Purpose: Introduce the characters, the setting, and the first wing (The Gateway). Establish the contrast between Elias’s intellectualism and Maya’s influencer persona.
- Emotional Arc: From confident posturing and professional detachment to the very first hint of physical vulnerability (the bead of sweat).
- Est. Words: 600
Key Events: { “introduction” : “Marcus Reed introduces the Gauntlet and the concept of culinary vs carnal.”, “character_posturing” : “Elias uses academic language (Baudrillard) while Maya focuses on her digital audience.”, “the_first_wing” : “The Gateway (50,000 Scoville) is served.”, “reaction” : “Elias dismisses the heat as ‘pedestrian’ while Maya embraces the ‘vibe’.”, “foreshadowing” : “A single bead of sweat appears on Elias’s hairline, signaling the start of the physical struggle.” }
Scene 2: The Terroir of Agony
- Setting: gauntlet_studio
- Characters: Elias Thorne, Maya Vance, Marcus “The Architect” Reed
- Purpose: Escalate the physical and psychological tension with the second wing (The Catalyst). Demonstrate the ‘Sophistication Paradox’.
- Emotional Arc: Increasing physical distress masked by intellectualization (Elias) and performative authenticity (Maya). The transition from control to the beginning of a breakdown.
- Est. Words: 750
Key Events: { “the_second_wing” : “Marcus introduces The Catalyst (250,000 Scoville).”, “intellectual_defense” : “Elias attempts to analyze the ‘neoliberal approach to spice’ while his voice falters.”, “social_friction” : “Maya mocks Elias’s physical state for her stream, highlighting his ‘beige’ appearance.”, “biological_betrayal” : “Marcus notes Elias’s dilated pupils; the body fights the mind’s narrative.”, “the_crack” : “Elias coughs and his eyes well up, breaking the clinical silence and the illusion of control.” }
Act 2: The Semiotic Collapse
Purpose: To dramatize the ‘Prisoner’s Dilemma’ of pain and the breakdown of the characters’ public personas.
Scene 1: The Lexical Erosion
- Setting: gauntlet_studio
- Characters: Elias Thorne, Maya Vance, Marcus “The Architect” Reed
- Purpose: To depict the transition from ‘spicy food’ to ‘chemical weapon’ and the first major victory of the body over the mind.
- Emotional Arc: From attempted sophistication and performance to raw biological panic and weaponized suffering.
- Est. Words: 1000
Key Events: { “1” : “Introduction of ‘The Obsidian Needle’ (850,000 SHU) and the shift in studio atmosphere to high-contrast violet.”, “2” : “Elias attempts an intellectual monologue but suffers a total loss of eloquence as the heat hits, failing to finish the word ‘palimpsest’.”, “3” : “Maya transitions from traditional performance to weaponizing her physical distress for the camera, claiming her suffering is ‘the real me’.”, “4” : “Marcus taunts Elias about the ‘colony in revolt,’ highlighting the shift from the semiotic to the somatic.” }
Scene 2: The Red Queen Race
- Setting: gauntlet_studio
- Characters: Elias Thorne, Maya Vance, Marcus “The Architect” Reed
- Purpose: To demonstrate the ‘Prisoner’s Dilemma’ of pain and the total collapse of persona into organism.
- Emotional Arc: From competitive dread to existential biological survival; mutual degradation and the death of the persona.
- Est. Words: 1200
Key Events: { “1” : “Introduction of ‘The Singularity’ (1.6 Million SHU) and the establishment of the ‘Prisoner’s Dilemma’ where neither participant wants to be the first to quit.”, “2” : “Maya experiences a total persona collapse, emitting a biological ‘error code’ instead of speech after taking a bite.”, “3” : “Elias’s pride forces him to continue despite his brain processing only the signal of ‘Fire’, losing all ability to formulate industrial critiques.”, “4” : “Marcus observes the ‘birth of the organism’ as the characters reach a physical and psychological stalemate, staring each other down over the milk.” }
Act 3: The Altar of the Scoville
Purpose: To reach the ‘Threshold of Semiotic Collapse’ and resolve the conflict through a moment of ‘Biological Honesty.’
Scene 1: The Singularity
- Setting: gauntlet_studio
- Characters: Elias Thorne, Maya Vance, Marcus “The Architect” Reed
- Purpose: This scene depicts the consumption of the final wing, representing the total breakdown of the characters’ carefully constructed personas.
- Emotional Arc: The characters transition from performative sophistication and calculated energy to a state of primal, mammalian dread and total chemical emergency.
- Est. Words: 750
Key Events: { “introduction” : “Marcus introduces ‘The Singularity’, a wing coated in six million Scoville units of capsaicin extract.”, “consumption” : “Elias and Maya consume the final wing, leading to an immediate breakdown of their personas.”, “collapse” : “The characters experience a total semiotic collapse, where language and branding dissolve into raw biological reaction.” }
Scene 2: The Cooling Down
- Setting: gauntlet_studio
- Characters: Elias Thorne, Maya Vance, Marcus “The Architect” Reed
- Purpose: The aftermath of the physical pain where artifice is stripped away and a moment of genuine, unmediated human connection occurs between the two rivals.
- Emotional Arc: A transition from intense physical suffering to a quiet, shared vulnerability and genuine human connection.
- Est. Words: 500
Key Events: { “production_ends” : “Marcus calls ‘cut’ and the theatrical studio lights are replaced by soft, mundane amber lighting.”, “recovery” : “Elias and Maya begin to recover, speaking to each other without their usual masks of sophistication or engagement.”, “connection” : “The rivals find common ground in the shared trauma of their experience, achieving ‘Biological Honesty.’”, “conclusion” : “The characters are led off-set, leaving behind a messy studio that reflects the end of the performance.” }
Status: ✅ Complete
Setting: gauntlet_studio
Prompt:

Character: Elias Thorne
Prompt:

Character: Maya Vance
Prompt:

Character: Marcus “The Architect” Reed
Prompt:

## The Semiotic Appetizer
Act 1, Scene 1
Setting: gauntlet_studio
Characters: Elias Thorne, Maya Vance, Marcus “The Architect” Reed
The studio was a vacuum of polished obsidian and surgical steel, a sterile stage bathed in the unforgiving halo of ring lights. At the center sat a triangular table, its surface so reflective that Elias Thorne could see the distorted underside of his own jaw. Three high-definition cameras tracked him with the silent, predatory hunger of deep-sea creatures.
To his left, Maya Vance performed a ritual of digital vanity. She tilted her head, checking her reflection in the lens of Camera Two, smoothing a stray hair into her honey-blonde blowout. She wasn’t looking at the room; she was looking at the imaginary millions she carried in her pocket like a digital rosary.
“Welcome to the Scoville Gauntlet,” Marcus Reed announced. His baritone was cultivated and clinical, a scalpel of sound. Known as ‘The Architect,’ Marcus stood at the head of the table, his charcoal suit as sharp as the lighting. “Tonight, we move beyond the artifice of the palate. We are not here for the culinary, but for the carnal. We are here to see what remains of the persona when biology takes over.”
Elias offered a thin, practiced smile. “A fascinating premise, Marcus. Though I suspect your ‘carnal’ is merely another layer of performance. We’re participating in a Baudrillardian hyperreality—the pain is real, yes, but the display of pain is the only currency that matters here.”
Maya let out a melodic, hollow laugh. “Elias, babe, you’re overthinking it. It’s just a vibe. People want to see us be real. They want the raw, unedited journey. Right, guys?” She winked at her primary camera, her thumb hovering over an invisible ‘post’ button.
“Then let the journey begin,” Marcus said. A masked assistant placed two white porcelain plates on the table. On each sat a single chicken wing, coated in a deceptive, sunset-orange glaze.
“The Gateway,” Marcus declared. “Thai Bird’s Eye chilies with a fermented habanero base. Fifty thousand Scoville units. A mere greeting.”
Elias picked up the wing with silver tongs, refusing to soil his fingers. He examined it with the detached curiosity of a pathologist. “Fifty thousand. Pedestrian. The heat equivalent of a brisk autumn breeze. A minor irritant to the mucous membranes, easily suppressed by the intellect.”
Maya used her manicured fingers, posing as the sauce glistened under the LEDs. “It smells… spicy-sweet. Like a bad breakup.” She took a dainty, deliberate bite. Her eyes widened for the lens. “Oh, wow. Okay. That’s a kick. It’s like a sparkler going off on my tongue.”
Elias took a bite. The heat was immediate—a sharp, needle-like prickling across his soft palate. He chewed slowly, maintaining a mask of scholarly indifference as the warmth slid down his esophagus like a swallowed coal.
“The capsaicin binds to the TRPV1 receptors,” Elias said, his voice steady despite a tightening throat. “The brain receives a signal of thermal abrasion, yet there is no tissue damage. It is the ultimate lie of the nervous system.”
“Is it a lie if it hurts?” Maya asked, her breath hitching. Her cheeks were flushing a cinematic pink. “I feel so… awake. My heart is actually racing.”
“Adrenaline,” Elias dismissed, though his own pulse throbbed against his collar. He reached for his water, then stopped. He would not be the first to break.
He looked at Marcus, ready to deliver another critique, but the words snagged. A single, microscopic bead of moisture had formed at the center of Elias’s hairline. A salt-rimmed traitor. He felt it—a cold prickle against his skin that felt like a mountain of ice in the desert of his burning mouth. The Gateway was open, and the biological reality was already eroding the social artifice.
Word Count: 610
Key Moments:
- Marcus Reed sets the philosophical and physical stakes of the Gauntlet, framing it as a battle between persona and biology.
- Elias attempts to intellectualize the experience using academic theory (Baudrillard), while Maya frames it as an ‘authentic’ moment for her digital brand.
- The first wing, ‘The Gateway’ (50,000 SHU), is consumed, serving as the baseline for the physical escalation to come.
- Despite his verbal dismissal of the heat, Elias experiences the first involuntary physical symptom—a bead of sweat—signaling the beginning of his loss of control.
Character States:
- Elias Thorne: Emotionally arrogant and intellectually detached; physically, his body is beginning to react to the capsaicin despite his mental resistance (the bead of sweat).
- Maya Vance: Emotionally performative and focused on her ‘brand’; physically, she is experiencing a ‘high’ from the adrenaline and heat, leaning into the sensation for the camera.
- Marcus Reed: Calm, authoritative, and observant; he remains the detached ‘Architect’ overseeing the experiment.
Status: ✅ Complete
Act 1, Scene 1 Image
Prompt:

## The Terroir of Agony
Act 1, Scene 2
Setting: gauntlet_studio
Characters: Elias Thorne, Maya Vance, Marcus “The Architect” Reed
The bead of sweat on Elias Thorne’s forehead was no longer a solitary scout; it had been joined by a battalion. The studio lights, once merely bright, now functioned like heat lamps in a vivarium. Across the obsidian table, Marcus “The Architect” Reed sat with the stillness of a gargoyle, his gloved hands hovering over a tray of porcelain ramekins.
“The Gateway was a courtesy,” Marcus said, his voice a low, resonant cello. “A handshake. Now, we move to the conversation. This is ‘The Catalyst.’ Two hundred and fifty thousand Scoville units. A blend of Habanero and Scotch Bonnet, fortified with a distilled oleoresin.”
He placed the second wing before them. It was a deep, bruised orange, glistening with an oil that seemed to vibrate under the LEDs.
Maya Vance didn’t hesitate. She adjusted her ring light, her smile flashing with the practiced precision of a predator. “Look at that color, guys,” she whispered to her phone, her voice a breathy, curated intimacy. “It’s like a sunset in a bottle. Or a warning. Elias, you look a little… muted. Are we losing our color?”
Elias felt the heat from the first wing radiating in his gut, a slow-blooming nebula of discomfort. He adjusted his glasses, which were beginning to slide down the bridge of his nose.
“Color is a bourgeois preoccupation, Maya,” Elias said, though his voice lacked its usual resonance. It sounded thin, like parchment being stretched. “What we’re seeing here is the neoliberal approach to spice. It’s not about flavor; it’s about the performative endurance of a manufactured crisis. We’ve commodified the biological fight-or-flight response into a consumable spectacle.”
He picked up the wing. His fingers betrayed a micro-tremor he hoped the cameras wouldn’t catch. He took a bite.
The heat was immediate and vertical. It didn’t wash over his tongue; it pierced it. It was a chemical needle, threading through his mucous membranes and stitching his throat shut. The Catalyst lived up to its name, igniting a chain reaction that turned his saliva into molten lead.
“Interesting,” Marcus noted, leaning forward. He wasn’t looking at the wings; he was studying Elias’s eyes. “The Sophistication Paradox in real-time. The more the mind attempts to categorize the pain, the more the body rebels against the categorization. Elias, your pupils are blown. Mydriasis. Your sympathetic nervous system has decided that Baudrillard can’t save you from a capsaicin burn.”
Maya laughed, a sharp, bright sound. She had taken her bite and was leaning into the burn, her face flushed a vibrant, healthy pink. “He’s turning beige, Marcus. Is beige a color? He looks like a library book that’s been left in the sun.” She turned her camera toward Elias, capturing the sheen of moisture on his upper lip. “Elias, tell the fans about the… what was it? The manufactured crisis? You look like you’re having a real one.”
Elias tried to retort. He had a brilliant point about the intersection of sensory overload and the erosion of the ego, but the words were trapped behind a wall of fire. His throat felt as though it had been lined with crushed glass. The “terroir” Marcus had promised was a landscape of scorched earth.
He swallowed, and the movement felt like a betrayal. The heat descended into his esophagus, a slow-moving lava flow.
“The… the irony,” Elias managed, his voice cracking—a jagged sound that broke the clinical silence of the room. “The irony is that the… the body’s insistence on reality… is the only thing… that isn’t… simulated.”
A sudden, violent tick developed in his left eyelid. His vision blurred as his tear ducts, sensing the chemical assault, began to overcompensate. A single, heavy tear escaped, carving a path through the fine dusting of translucent powder on his cheek.
Then came the cough.
It was a dry, hacking sound—the sound of a man trying to expel his own lungs. Elias clutched the edge of the table, his knuckles white. The intellectual defense had failed. The fortress of his vocabulary had been breached by a pepper.
“The crack,” Marcus whispered, almost to himself. He looked satisfied, like a scientist watching a crystal lattice finally shatter under pressure. “The social artifice is melting, Elias. Tell me, in this moment, does the ‘neoliberal approach’ matter? Or is there only the burn?”
Elias couldn’t answer. He reached for his glass of water, his hand shaking so violently that the ice clinked a frantic rhythm against the glass. He stopped just short of drinking, his pride a dying ember in the furnace of his mouth. He looked at Maya, who was grinning, her eyes wide and hungry for the content of his collapse.
He forced himself to set the glass down. His eyes were bloodshot, swimming in involuntary salt water, but he stared at Marcus with a raw, primal defiance.
The Catalyst had done its work. The conversation had truly begun.
Word Count: 813
Key Moments:
- Marcus escalates the heat to 250,000 SHU with ‘The Catalyst’, moving from a ‘handshake’ to a ‘conversation’.
- Elias attempts to frame the pain as a neoliberal construct, but his physical symptoms (dilated pupils, trembling) betray his mental detachment.
- Elias suffers a coughing fit and an involuntary tear, marking the first major crack in his composed, academic persona while Maya weaponizes his distress for her audience.
Character States:
- Elias Thorne: Physically distressed and losing control; his throat is ‘scorched earth,’ and his eyes are tearing up. Emotionally, he is desperate to maintain his intellectual superiority but is failing.
- Maya Vance: Physically flushed and ‘high’ on the adrenaline; she is leaning into the pain to appear ‘authentic’ and is thriving on Elias’s visible struggle.
- Marcus Reed: Calm, clinical, and observant; he is pleased to see the ‘Sophistication Paradox’ taking effect and the breakdown of Elias’s social mask.
Status: ✅ Complete
Act 1, Scene 2 Image
Prompt:

## The Lexical Erosion
Act 2, Scene 1
Setting: gauntlet_studio
Characters: Elias Thorne, Maya Vance, Marcus “The Architect” Reed
The studio lights pivoted with a hydraulic hiss. The inviting amber of the previous round vanished, replaced by a bruised, high-contrast violet that turned the sweat on Elias’s brow into beads of cold mercury. The air conditioning hummed—a low-frequency vibration that didn’t cool the room so much as rattle the marrow of his teeth.
Marcus Reed, “The Architect,” sat behind the obsidian table like a judge presiding over a neon purgatory. Between them lay the next offering: a single chicken wing coated in a sauce so dark it seemed to absorb the violet light, a matte void on a white ceramic plate.
“The Obsidian Needle,” Marcus announced, his voice a smooth, terrifying baritone. “Eight hundred and fifty thousand Scoville Heat Units. We have moved past the ‘Catalyst.’ We are now entering the realm of the chemical weapon. This is no longer a culinary experience, Elias. It is a biological intervention.”
Elias Thorne stared at the wing. His mouth was already a ruin, a landscape of scorched earth where his taste buds used to reside. His pulse thrummed in his ears, a frantic, syncopated rhythm. He felt the phantom weight of his reputation—decades of sharp-tongued critiques and the effortless deconstruction of culture—pressing down on his shoulders. He needed to speak. He needed to colonize this pain with language before it colonized him.
“It’s… a fascinating escalation,” Elias said, his voice cracking like dry parchment. He cleared his throat, the sound wet and jagged. “The transition from the gustatory to the purely… purely somatic. It’s a classic postmodern maneuver. We are stripping away the signifier of ‘food’ to reveal the raw, unmediated… the raw…”
He reached for the wing. His fingers trembled, a fine-motor betrayal he couldn’t suppress. He took a bite.
For three seconds, there was nothing but the metallic tang of vinegar. Then, the Needle struck.
It wasn’t heat. Heat was a campfire; heat was a summer day. This was a localized tectonic shift. It felt as though a thousand microscopic glass shards had been dipped in battery acid and driven into the soft tissue of his throat. His sinuses didn’t just open; they screamed.
“The beauty of this,” Elias gasped, his eyes bulging, “is the way it… it functions as a… a palim—”
The word died. Palimpsest. He wanted to say palimpsest—to describe how the pain was overwriting his previous sensory experiences. But the ‘p’ was a puff of hot air, and the ‘l’ was a strangled sob. His tongue had become a foreign object, a heavy, swollen slug that refused to obey the commands of his prefrontal cortex. He gripped the edge of the table, his knuckles white. The violet light pulsed in time with the throbbing in his carotid artery.
Beside him, Maya Vance was undergoing a different kind of transformation.
She wasn’t fighting the erosion; she was curating it. Tears streamed down her face, carving tracks through her high-definition foundation, but she didn’t wipe them away. She leaned into the camera, her breath coming in ragged, shallow hitches. Her eyes, bloodshot and shimmering, were fixed on the lens with a terrifying, predatory intensity.
“Do you see this?” she whispered, her voice a raspy ghost of its usual melodic lilt. She held up her shaking hand, the Obsidian Needle held like a grim trophy. “This isn’t a filter. This isn’t a script. This is… this is the real me. I’m burning. I’m literally burning for you guys.”
She took another bite, a deliberate act of self-immolation. She winced, a spasm of genuine agony crossing her face, but she immediately smoothed it into a look of tragic ecstasy.
“Maya,” Marcus said, leaning forward, “you’re weaponizing your own distress. It’s brilliant. You’ve turned a biological crisis into a brand identity.”
“It’s… authenticity,” Maya choked out. A string of saliva escaped the corner of her mouth, but she didn’t flinch. The engagement metrics in her head were surely skyrocketing. “Pain is the only thing… the only thing they can’t… fake.”
Marcus turned his gaze back to Elias, who was currently hunched over, his face a deep, alarming shade of plum. Elias was trying to breathe through his nose, but his lungs felt like they were filled with hot sand.
“How’s the monologue coming, Elias?” Marcus asked, his tone dripping with clinical curiosity. “You look like a man who has lost his dictionary.”
Elias looked up. He tried to summon a scathing retort, a witty observation about the banality of Marcus’s sadism. He opened his mouth, and for a moment, the ghost of a sentence formed in his mind: The architectural cruelty of this environment suggests a—
“Guh,” Elias said.
That was it. A guttural, primal sound. The “colony” of his body had officially declared independence from the “governor” of his mind. His nervous system was screaming so loudly that the intellect had been evicted.
“The colony is in revolt,” Marcus said, echoing Elias’s internal thought with haunting accuracy. “The semiotic has been conquered by the somatic. You wanted to talk about the ‘social artifice’ of the Gauntlet, Elias. But right now, you aren’t a critic. You aren’t a scholar. You’re just a collection of nerve endings trying not to vomit.”
Elias felt a tear escape his left eye. It wasn’t a tear of sadness; it was a tear of pure, chemical exhaustion. He looked at the glass of milk sitting just out of reach. It looked like a holy relic, a white beacon of salvation. His pride, which had felt like an iron suit of armor only ten minutes ago, was now a lead weight dragging him into the abyss.
Maya let out a sharp, jagged laugh that turned into a cough. “He’s… he’s broken,” she wheezed, pointing a trembling finger at Elias. She looked at the camera, her expression one of manic triumph. “The Great Elias Thorne… out of words. I guess… I guess I’m the only one… still talking.”
She was right. She was winning because she had accepted the degradation. She had traded her dignity for “content,” while Elias was still trying to trade his pain for “meaning.” And in the violet glow of the Obsidian Needle, meaning was a currency that no longer held any value.
Elias reached out. His hand hovered over the milk. He looked at Marcus, whose eyes were cold and expectant. He looked at Maya, who was filming his defeat with a predatory grin.
He didn’t drink. Not yet. But the silence where his words used to be was the loudest sound in the room.
“Next round,” Marcus said, his voice a death knell. “We go to one point two million. Let’s see if there’s anything left of you to burn.”
Word Count: 1109
Key Moments:
- The Introduction of the Obsidian Needle: The stakes are raised to 850,000 SHU, and the studio environment shifts to a hostile, high-contrast violet, signaling the move from ‘food’ to ‘chemical weapon’.
- The Lexical Collapse: Elias attempts to maintain his intellectual superiority through a monologue but suffers a total breakdown of language, failing to complete the word ‘palimpsest’ as his biology overrides his intellect.
- Maya’s Weaponized Suffering: Maya leans into her physical distress, using her tears and pain to claim a new level of ‘authenticity’ for her brand, effectively winning the social battle by embracing the biological one.
- The Architect’s Taunt: Marcus highlights the ‘colony in revolt,’ mocking Elias for losing his vocabulary and pointing out that the body has officially conquered the mind.
Character States:
- Elias Thorne: Physically shattered and intellectually silenced. He is in a state of ‘lexical erosion,’ where his primary tool—language—has failed him. He is hovering on the edge of total surrender.
- Maya Vance: Physically in pain but emotionally empowered. She has successfully commodified her suffering and feels a sense of triumph over Elias’s collapse.
- Marcus Reed: Calm, observant, and increasingly dominant. He is enjoying the ‘Sophistication Paradox’ as he watches the high-level thinkers and performers reduced to primal reactions.
Status: ✅ Complete
Act 2, Scene 1 Image
Prompt:

## The Red Queen Race
Act 2, Scene 2
Setting: gauntlet_studio
Characters: Elias Thorne, Maya Vance, Marcus “The Architect” Reed
The studio’s violet light had curdled into a bruised, ultraviolet rot, turning the white porcelain spoons into glowing, irradiated shards. The air was no longer oxygen; it was a saturated aerosol of pulverized peppers—a fine mist of capsaicin that clung to the soft tissue of the throat like a threat.
Marcus “The Architect” Reed sat in the center of the gloom, a sharp, ink-black silhouette against a wall of glowing monitors. Between Elias and Maya sat a single obsidian ramekin. Inside was a substance so dark it seemed to exert its own gravity—a viscous, maroon sludge that smelled less like food and more like an electrical fire in a chemical plant.
“This,” Marcus said, his voice dropping into a reverent, clinical register, “is The Singularity. One point six million Scoville Heat Units. We have moved past the threshold of culinary experience. We are now in the realm of neuro-chemical warfare.” He leaned forward, his eyes unblinking in the harsh light. “Your bodies will tell you that you are dying. Your brains will attempt to shut down non-essential systems to preserve the core. The question is: which of you is more than just a collection of panicked nerves?”
Elias Thorne stared at the sludge. His hands were trembling—a fine, high-frequency vibration he couldn’t suppress. His throat felt as though it had been lined with fiberglass. He wanted to speak, to offer a pithy observation about the fetishization of extremity in late-stage capitalism, but the words felt like dry sand in his mouth. He looked at Maya.
Maya Vance was no longer the polished avatar of lifestyle perfection. Her mascara had dissolved into dark, jagged canyons down her cheeks. Her breathing was shallow, ragged, the hyperventilation of a prey animal. Yet, she held her phone with a white-knuckled, skeletal grip. The “Live” icon was a pulsing red eye, a digital witness to her unraveling.
“The Prisoner’s Dilemma,” Marcus whispered. “The milk is right there. Cold, soothing, merciful. If one of you reaches for it, the other wins the round by default. You can end this. You can both end this. But the first one to move is the one who admits their persona is a lie. The first one to drink is the one who confesses they are merely an animal.”
Maya reached for her spoon first. It wasn’t a gesture of courage; it was a twitch of pure, reflexive desperation. Elias followed, his pride a heavy, cold stone in his gut.
They took the dose simultaneously.
For three seconds, there was a deceptive, mocking silence. Then, the Singularity hit.
It wasn’t heat. Heat was a domestic concept, something associated with hearths and comfort. This was a localized supernova. It was as if a molten needle had been driven through the center of the tongue and out the back of the skull.
Maya’s reaction was immediate and terrifying. She didn’t scream; she didn’t have the breath for it. Her body suffered a total system failure. Her eyes rolled back, showing only the bloodshot whites, and her jaw began to work in a rhythmic, mechanical fashion. A sound escaped her—a high-pitched, staccato clicking of teeth against teeth that sounded like a corrupted audio file. It was a biological error code, the sound of a sophisticated social interface crashing under the weight of raw, unmediated agony. She dropped her phone. The screen shattered, but the camera remained upward, filming the ceiling, capturing the distorted, violet-tinged reality of her collapse.
Elias felt his own mind begin to fracture. He tried to cling to his intellect, to find the vocabulary to describe the sensation. The semiotics of the sear, he thought. The ontological erasure of the self through—
The thought died. The word fire rose up like a monolith, obliterating everything else. His brain, once a library of complex theories and sharp-edged critiques, was now a burning room with only one exit, and that exit was locked.
Fire.
He looked at the milk. It was a white beacon of salvation, a liquid ghost. His hand moved toward it, an inch, then stopped. He looked at Maya. She was slumped over the table, her forehead resting on the cool wood, that horrific clicking sound still emanating from her throat.
If he drank, she won. If he drank, the “Cultural Critic” was just a man who couldn’t handle a pepper. He would be a footnote in her next viral apology video. He would be the man who broke first.
Fire. Fire. Fire.
The pain moved from his mouth to his ears, a deafening roar of static. His vision began to tunnel, the violet light turning black at the edges. He could feel his sweat—cold, oily, and smelling of capsaicin—soaking through his bespoke linen shirt. He was no longer Elias Thorne, the man who had dismantled the pretensions of the Venice Biennale. He was a lung, a heart, and a screaming nervous system. He was a primate in a suit, trapped in a cage of his own making.
Marcus watched them with the detached fascination of an entomologist observing a pinned specimen. “Look at you,” he murmured. “The influencer and the intellectual. Reduced to the fundamental state. The organism has finally evicted the ego. There is no brand here, Maya. There is no discourse here, Elias. There is only the red.”
Elias stared at Maya. Through the haze of his dissolving consciousness, he saw her eyes snap back into focus. They were bloodshot, leaking fluid, and filled with a primal, predatory hate. She wasn’t looking at him as a rival; she was looking at him as an obstacle to her survival.
She reached for the milk.
Elias’s hand shot out, not to grab his own glass, but to hover near hers. It was a silent, agonizing stalemate. If she moved, he would move. They were locked in a Red Queen race—running as hard as they could just to stay in the same place of absolute, unendurable suffering.
Neither would drink. Neither would yield.
The “Prisoner’s Dilemma” had reached its terminal phase. To save themselves was to lose everything they had built their lives upon. To maintain the lie was to endure a physical hell that their bodies were not designed to survive.
Maya’s hand trembled violently, her fingers inches from the glass. A single drop of saliva escaped her lips, hitting the table like a heavy tear. She looked at Elias, and for a brief, flickering second, the masks were gone. There was no “Vance Lifestyle,” no “Thorne Critique.” There were just two drowning people, each holding the other’s head under the water to keep themselves afloat.
“The birth of the organism,” Marcus said, his voice sounding like it was coming from the end of a long, dark tunnel. “Beautiful. You’ve stopped pretending to be people. You’re finally being honest.”
Elias tried to form a word. He wanted to say enough. He wanted to say mercy.
But his tongue was a swollen, dead weight. His brain sent the signal, but the hardware was fried. The only thing left was the signal of the fire, a constant, unwavering scream in the dark.
He leaned forward, his face inches from Maya’s. Their breath, hot and smelling of vinegar and destruction, mingled in the narrow space between them. They were a tableau of shared degradation, a monument to the vanity of the human will.
“Round three,” Marcus announced, his voice cutting through the static of their pain like a blade. “The Finality. Two point two million. If you’re still alive, that is.”
Elias didn’t move. Maya didn’t move. They remained frozen, staring at each other over the white glasses of milk, two ghosts waiting for the fire to finish what it had started.
Word Count: 1281
Key Moments:
- Marcus introduces ‘The Singularity’ (1.6 Million SHU), framing the round as a ‘Prisoner’s Dilemma’ where the first to seek relief loses social standing.
- Maya suffers a total biological breakdown, losing the ability to speak and emitting a ‘biological error code’ as her persona is erased by pain.
- Elias’s internal monologue is reduced to the single word ‘Fire,’ as his brain loses the capacity for complex thought or cultural critique.
- The scene ends in a physical and psychological deadlock, with both characters hovering over the milk but refusing to drink, locked in a mutual struggle for survival and pride.
Character States:
- Elias Thorne: Physically devastated and intellectually hollowed out. He is functioning on pure, stubborn pride, his mind having devolved from a complex analytical tool to a simple receiver for the signal of ‘Fire.’
- Maya Vance: Physically broken and socially ‘glitched.’ Her carefully curated image has been replaced by a raw, animalistic survival instinct. She is in a state of ‘biological error,’ unable to maintain her performance.
- Marcus Reed: Dominant, clinical, and satisfied. He has successfully stripped away the social artifice of his guests and is relishing the ‘birth of the organism’ as they reach their breaking points.
Status: ✅ Complete
Act 2, Scene 2 Image
Prompt:

## The Singularity
Act 3, Scene 1
Setting: gauntlet_studio
Characters: Elias Thorne, Maya Vance, Marcus “The Architect” Reed
The studio air had curdled. It was no longer a mix of oxygen and nitrogen, but a suspended aerosol of vinegar, capsaicin, and the metallic tang of adrenaline. The violet lights of the previous round had deepened into a bruised, necrotic crimson that pulsed in time with the throbbing in Elias’s temples.
Marcus “The Architect” Reed reached beneath the table. He moved with the terrifying precision of a surgeon. When he placed the final tray on the obsidian surface, the sound of ceramic hitting stone carried the finality of a gunshot.
“The Singularity,” Marcus whispered. His voice was the only thing in the room that remained structural. “Six million Scoville units. This is not a condiment; it is a chemical coup d’état. It does not interact with your palate; it colonizes your nervous system. At this concentration, the brain ceases to interpret ‘flavor’ and begins to signal ‘systemic failure.’”
On the tray sat two wings, coated in a paste so dark it appeared to absorb the studio lights. It looked like cooled lava, or perhaps the charred remains of a language.
Elias Thorne stared at the meat. His mind, once a library of meticulously filed critiques and polysyllabic barbs, was now scorched earth. He tried to summon a word—transcendence, nihilism, visceral—but the folders were empty. He was a man who had built a career on the mediation of experience through prose, yet he was now facing an experience that refused to be mediated.
To his left, Maya Vance was a ruin of curated beauty. Mascara tracked black canyons down her cheeks, and her breathing was a wet, rhythmic hitching. The “glow” she had marketed to millions had fermented into a sickly, feverish sheen. She wasn’t looking at the camera anymore. The camera was a dead eye. The only thing that existed was the black mass on the plate.
“To the end of the self,” Marcus said, a thin, cruel smile touching his lips.
They picked up the wings. Elias’s fingers trembled so violently the bone-dry skin of the chicken rattled against the plate. He looked at Maya. For a fleeting second, the social artifice flickered—the critic and the influencer, the high-brow and the low-brow—and then it vanished. They were just two mammals in a trap.
They bit.
The collapse was instantaneous.
For Elias, the world didn’t catch fire; it folded. The six million units hit his tongue not as heat, but as a physical weight—a crushing, tectonic pressure that flattened his tongue and surged into his sinuses like molten lead. His vision fractured. The studio walls drifted away, replaced by a screaming, white-hot static. He tried to scream, but his throat had seized, the muscles locking in a desperate, futile attempt to wall off the poison.
I am… he thought. But the sentence wouldn’t finish. The ‘I’ was being erased. There was no Elias Thorne. There was only the Signal. The Signal was red. The Signal was agony. The Signal was the biological reality of a body realizing it had been betrayed by the mind.
Maya collapsed forward, her forehead hitting the table with a dull thud. She wasn’t crying anymore; she was leaking. Saliva, tears, and sweat pooled on the black surface, reflecting the red lights above. Her brand, her “authentic” journey, her carefully managed vulnerability—it had all been incinerated. She let out a sound that was neither human nor animal, a low, guttural vibration that bypassed the vocal cords and emerged directly from the lungs. It was a biological error code.
Marcus leaned back, his silhouette sharp against the crimson haze. He watched them with clinical detachment.
“Look at you,” Marcus said, his voice sounding as if it were coming from the end of a long tunnel. “The critic has no words. The idol has no image. You’ve reached the Singularity. The point where the persona is crushed by the weight of the organism.”
Elias tried to lift his head. He wanted to retort, to find one last shred of intellectual dignity, but his brain had decoupled from his jaw. He looked at the glass of milk—the white flag of surrender—but his hand wouldn’t move. He was paralyzed in a state of pure, unadulterated sensation.
The semiotic collapse was total. The word ‘wing’ meant nothing. The word ‘show’ meant nothing. There was only the fire, the rhythmic thud of a heart trying to outrun a ghost, and the terrifying realization that beneath the clothes and the titles, there was nothing but a shivering, panicked animal.
They sat in the red dark, two ghosts of the people they had been five minutes ago, waiting for the world to start again.
Word Count: 771
Key Moments:
- The Introduction of The Singularity: Marcus presents the final wing, a 6-million SHU extract-heavy nightmare, framing it as the end of the “self.”
- The Final Bite: Elias and Maya consume the wing, triggering an immediate and violent biological override of their consciousness.
- The Total Collapse: Both characters lose the ability to maintain their personas; Elias loses language entirely, and Maya’s physical breakdown destroys her aesthetic brand.
Character States:
- Elias Thorne: Physically paralyzed and intellectually void. His internal monologue has shattered into sensory fragments; he is no longer a critic, but a “shivering animal.”
- Maya Vance: Physically broken and socially erased. She has moved past “performative suffering” into a state of genuine chemical emergency, her image completely destroyed by the raw reality of her body’s reaction.
- Marcus Reed: Ascendant and god-like. He has achieved his goal of stripping away the social artifice of his guests, observing their “biological error codes” with clinical satisfaction.
Status: ✅ Complete
Act 3, Scene 1 Image
Prompt:

## The Cooling Down
Act 3, Scene 2
Setting: gauntlet_studio
Characters: Elias Thorne, Maya Vance, Marcus “The Architect” Reed
The red glare of the studio lights died, replaced by the flat, apologetic amber of the overheads. It was a jarring eviction from the theater of pain back into the mundane reality of a Long Island City warehouse. The air, once thick with the scent of vinegar and adrenaline, now tasted of dust and industrial cooling.
“And we’re clear,” Marcus said. His voice, previously a booming toll, had flattened into the drone of a man checking a grocery list. He unclipped his lapel mic, eyes already fixed on a playback monitor. “Good stuff, guys. Truly. We’ll get the ‘after-action’ stills in ten.”
Elias felt the world rush back in through his pores. The “Fire” had retreated from a roar to a low, rhythmic throb, leaving his nervous system a scorched landscape. He reached for the glass of milk. There was no pride left to prevent the surrender, no audience to witness the defeat. He drank, the cold liquid coating his throat like liquid marble—a desperate masonry repair for his shattered dignity.
Across the table, Maya’s practiced grace had evaporated. A streak of mascara cut a dark canyon through her foundation. She looked at Elias, and for the first time, her eyes weren’t searching for a lens or a light source. They were wide, dark, and entirely human.
“My teeth,” she whispered, her voice a raspy ghost. “I think my teeth are vibrating.”
Elias set the glass down with a shaky hand. He tried to summon a biting remark about the commodification of suffering, but the words felt heavy and useless, like wet sand. “It’s the endorphin dump,” he managed, his voice cracking. “The brain is attempting to compensate for the trauma.”
Maya let out a sound that might have been a laugh if she had the breath for it. She leaned forward, resting her forehead against the cool surface of the table, heedless of the orange grease and discarded bones. “Trauma. Yeah. That’s a word for it.”
They sat in silence for a long minute, the only sound the hum of industrial fans and the distant chatter of the crew. In this quiet, the artifice of their rivalry—the critic versus the creator—evaporated. They weren’t brands; they were biology.
“You didn’t quit,” Elias said, surprised by the genuine respect in his tone.
Maya lifted her head, her face pale and glistening. “Neither did you, Professor. Even when you looked like you were seeing God.”
“I wasn’t seeing God,” Elias muttered, a faint smile touching his swollen lips. “I was seeing the heat death of the universe. It was surprisingly orange.”
A production assistant approached with damp towels. “Cooling tent, now. Please follow me.”
They rose together, their legs unsteady. As they were led off-set, Elias glanced back. The “Singularity” wings sat there, cold and innocuous, surrounded by crumpled napkins and spilled milk. The stage was a wreck, a messy testament to the collapse of their curated personas. They walked into the shadows, two ghosts bound by the shared, silent language of the burn.
Word Count: 502
Key Moments:
- The Shift: The theatrical red lights are replaced by mundane amber, signaling the end of the performance and the return to reality.
- The Recovery: Elias and Maya finally drink the milk, abandoning their pride for the sake of physical relief.
- Biological Honesty: The rivals share a moment of genuine, unscripted connection, acknowledging their shared trauma without the masks of their public personas.
Character States:
- Elias Thorne: Physically exhausted and humbled. His intellectual defenses have been stripped away, replaced by a raw, quiet respect for his opponent.
- Maya Vance: Disheveled and physically broken, but mentally clear. She has abandoned her “aesthetic” for the first time, finding a strange peace in the wreckage of her image.
- Marcus Reed: Clinical and detached. Having extracted the “truth” from his guests, he is already moving on to the logistics of production, treating the human subjects as mere data points.
Status: ✅ Complete
Act 3, Scene 2 Image
Prompt:

Final Statistics
- Total Scenes: 6
- Total Word Count: 5086
- Time: 712.914s
Game Theory Analysis
Started: 2026-02-20 20:08:39
Game Theory Analysis
Scenario: The Spicy Food Semiotic Field: A signaling game where the Consumer attempts to signal toughness or authenticity through the consumption of extreme heat, and the Observer interprets these signals to grant or withhold social status. Players: Consumer, Observer
Game Type: non-cooperative
Game Structure Analysis
This analysis explores the Spicy Food Semiotic Field through the lens of game theory, specifically as a Signaling Game with asymmetric information and costly signals.
1. Identify the Game Structure
- Game Type: Non-cooperative Signaling Game. It is a dynamic game of asymmetric information where one player (the Sender/Consumer) sends a signal to another player (the Receiver/Observer).
- Timing: Sequential. The Consumer moves first by choosing a heat level; the Observer moves second by choosing a social response.
- Information: Imperfect and Asymmetric.
- The Consumer knows their “Type” (their actual biological tolerance for capsaicin).
- The Observer cannot see the Consumer’s internal pain or tolerance directly; they only see the choice of food and the resulting physiological “Biological Honesty.”
- Repetition: Can be One-Shot (a single meal with strangers) or Repeated (social dynamics within a friend group or subculture), where reputation and “Social Capital” accumulate over time.
- Sum: Non-constant sum. The total utility can increase (both enjoy the “ritual of suffering”) or decrease (the Consumer is in agony and the Observer is unimpressed).
2. Define Strategy Spaces
Consumer (Sender) Strategies ($S_C$):
- Consume Extreme Heat ($s_{C1}$): High Pain / High Signal. An attempt to signal “Toughness” or “Authenticity.”
- Consume Mild/Moderate ($s_{C2}$): Low Pain / Low Signal. An attempt to prioritize comfort or avoid the risk of a failed performance.
- Constraints: Biological Honesty. The Consumer’s strategy is constrained by involuntary physiological responses (sweating, flushing). A “Low Tolerance” type attempting $s_{C1}$ may suffer a “leaky signal” that reveals their true type despite their strategic choice.
Observer (Receiver) Strategies ($S_O$):
- Grant Status ($s_{O1}$): Respect, validation, or inclusion in the “Chili-head” or “Authentic” subculture.
- Withhold Status ($s_{O2}$): Mockery, dismissal, or labeling the Consumer with the “Mayonnaise Proxy” (social sterility).
3. Characterize Payoffs
The payoffs are multi-dimensional, combining biological, psychological, and social factors.
Consumer Payoff ($U_C$):
\(U_C = (E + S) - (P + M + B)\)
- $E$ (Endorphin Loop): Private biological reward (positive).
- $S$ (Social Capital): Public status granted by the Observer (positive).
- $P$ (Physical Pain): The cost of capsaicin consumption (negative).
- $M$ (Mayonnaise Proxy): The social cost of choosing Mild in a high-stakes subculture (negative).
- $B$ (Bluffing Cost): The social penalty if “Biological Honesty” reveals the Consumer is faking toughness (negative).
Observer Payoff ($U_O$):
- (+) Social Cohesion: The benefit of a shared ritual or “suffering together.”
- (+) Information Accuracy: The utility of correctly identifying a “Tough” or “Authentic” peer.
- (-) Social Friction: The cost of awkwardness if the Consumer has a medical emergency or if the Observer’s mockery creates conflict.
| Consumer Choice | Observer: Grant Status | Observer: Withhold Status |
|---|---|---|
| Extreme Heat | High Social Capital, Endorphins, High Pain | High Pain, Mockery, Failed Performance |
| Mild/Moderate | Neutral Status, Low Pain | Mayonnaise Proxy, Low Pain, Social Dismissal |
4. Key Features & Strategic Dynamics
The Performance vs. Toughness Hypotheses
- Performance Hypothesis: The Consumer treats the meal as a Costly Signal. They incur the cost of pain ($P$) specifically to earn Social Capital ($S$). The signal is credible because it is painful.
- Toughness Hypothesis: The Observer uses the interaction to categorize the Consumer’s character. If the Consumer consumes heat without “Biological Honesty” (no sweat/tears), the Observer infers a “High Tolerance Type,” granting maximum status.
Information Asymmetry and Biological Honesty
The game is defined by the “Honest Signaling” principle. In nature, signals are honest if they are too expensive to fake. Here, the “Biological Honesty” (involuntary flushing) acts as a verification mechanism.
- If a Consumer chooses “Extreme Heat” but their body reacts violently, the Observer sees a “Separating Equilibrium”—the signal of toughness is undermined by the biological reality of weakness.
Equilibria States
- Separating Equilibrium: High-tolerance types eat spicy food to get status; low-tolerance types eat mild food to avoid pain. The signal accurately reflects the type.
- Pooling Equilibrium (The “Hot-Wing Industrial Complex”): Because the “Mayonnaise Proxy” (cost $M$) is so high in certain subcultures, everyone chooses Extreme Heat regardless of their tolerance. This leads to a “race to the bottom” where heat levels must continually escalate (Scoville-based marketing) to maintain the signal’s value.
The Sophistication Paradox
This represents a strategic contradiction. While the Consumer signals “Authenticity” through heat, the heat itself acts as a “Sensory Blackout Curtain,” destroying the ability to taste the food. The Consumer sacrifices the utility of flavor to gain the utility of status.
Payoff Matrix
This analysis explores the Spicy Food Semiotic Field through the lens of formal game theory, treating the consumption of capsaicin as a signaling game where biological pain is traded for social currency.
Part 1: Game Structure Analysis
1. Game Type
- Non-Cooperative: Players act in their own self-interest (Consumer seeks status/pleasure; Observer seeks accurate social categorization).
- Sequential Signaling Game: The Consumer moves first (chooses heat level); the Observer moves second (interprets the signal and chooses a social response).
- Asymmetric Information: The Consumer knows their true “type” (actual pain tolerance and internal physiological response), while the Observer only sees the outward “Biological Honesty” (sweat, redness) and the choice of heat.
2. Strategy Spaces
- Consumer ($S_C$):
- Extreme Heat ($S_H$): High-cost signal involving physical pain and potential “Biological Honesty” failure.
- Mild/Moderate ($S_M$): Low-cost signal, avoiding pain but risking the “Mayonnaise Proxy” penalty.
- Observer ($S_O$):
- Grant Status ($A_G$): Validation of the Consumer’s toughness or authenticity.
- Withhold Status ($A_W$): Mockery, dismissal, or labeling the Consumer as a “poser” or “bland.”
3. Payoff Components
- $P$ (Pain): The physical cost of capsaicin (Negative).
- $E$ (Endorphin Loop): The private biological reward following pain (Positive).
- $S$ (Social Capital): The public reward for a successful signal (Positive).
- $M$ (Mayonnaise Penalty): The social cost of choosing mildness in high-stakes subcultures (Negative).
- $B$ (Biological Honesty Constraint): If the Consumer chooses $S_H$ but their body reacts uncontrollably (tears, vomiting), the signal is “broken,” leading to a massive loss in $S$.
4. Key Features
- Costly Signaling: Extreme heat is a “Handicap Principle” signal. Because it hurts, it is difficult to fake, making it a potentially “honest” signal of toughness.
- The Performance Hypothesis: The game assumes the Consumer is “colonizing” their own pain to manipulate the Observer’s perception.
- The Toughness Hypothesis: The Observer uses the Consumer’s lack of reaction as a proxy for moral/physical character.
Part 2: Detailed Payoff Matrix
The following matrix represents a standard interaction within a “Chili-head” subculture or a high-stakes social setting (e.g., a first date at an authentic Thai restaurant).
Numerical Scale: -5 (Severe Loss) to +5 (Maximum Gain).
| Consumer Strategy \ Observer Action | Grant Status ($A_G$) | Withhold Status ($A_W$) |
|---|---|---|
| Consume Extreme Heat ($S_H$) | Outcome: The “Toughness” Success Payoffs: C: +3 / O: +2 |
Outcome: The “Try-Hard” Failure Payoffs: C: -2 / O: +1 |
| Consume Mild/Moderate ($S_M$) | Outcome: The “Authentic” Moderate Payoffs: C: +1 / O: +2 |
Outcome: The “Mayonnaise” Proxy Payoffs: C: -3 / O: +1 |
Breakdown of Payoffs:
1. Extreme Heat ($S_H$) + Grant Status ($A_G$)
- Consumer (+3): High Social Capital (+4) + Endorphin Loop (+2) - Physical Pain (-3). The Consumer successfully converted pain into status.
- Observer (+2): Gains the satisfaction of witnessing a “valid” performance and reinforces subculture norms.
- Logic: This is the Nash Equilibrium for a “Chili-head” subculture. Both players agree on the value of the ordeal.
2. Extreme Heat ($S_H$) + Withhold Status ($A_W$)
- Consumer (-2): Endorphin Loop (+2) - Physical Pain (-3) - Social Humiliation (-1). The Consumer endured the pain but was mocked as a “masochist” or “performer” rather than a “tough guy.”
- Observer (+1): Gains “Status Gatekeeper” utility. By withholding respect, the Observer asserts their own superior judgment or higher tolerance.
- Logic: This occurs when the Observer detects “Biological Honesty” failure (e.g., the Consumer is visibly suffering too much to be considered “tough”).
3. Mild/Moderate ($S_M$) + Grant Status ($A_G$)
- Consumer (+1): No Pain (0) + Social Comfort (+1). The Consumer is accepted as “sensible” or “authentic” without needing to perform.
- Observer (+2): Low effort required. The Observer appreciates the lack of drama and enjoys a peaceful meal.
- Logic: This represents a “Sophisticated” social circle where extreme heat is seen as gauche or destructive to flavor.
4. Mild/Moderate ($S_M$) + Withhold Status ($A_W$)
- Consumer (-3): No Pain (0) - Mayonnaise Proxy Penalty (-3). The Consumer is socially stigmatized for being “unadventurous” or “fragile.”
- Observer (+1): Gains utility from “Authenticity Policing.” By mocking the mild choice, they elevate their own status as someone who can handle the “real” version.
- Logic: This is the “Social Poison” of the semiotic field—where a simple dietary preference is weaponized as a character flaw.
Part 3: Strategic Observations
- The “Biological Honesty” Trap: If a Consumer chooses $S_H$ but has low genetic tolerance, the cost of $P$ (Pain) increases exponentially, and the probability of $A_W$ (Withholding Status) increases because the “performance” becomes visible. This creates a separating equilibrium where only those with high natural tolerance (or high desensitization) can profitably choose the Extreme Heat strategy.
- The Endorphin Loop as a Safety Net: The private payoff ($E$) ensures that even if the Observer withholds status, the Consumer may still find the game worth playing. This explains the “Chili-head” who eats alone; the game becomes a single-player interaction with their own nervous system.
- Pareto Efficiency: The (Mild, Grant Status) outcome is often the most Pareto-efficient (total utility is high with zero physical cost), but the “Toughness Hypothesis” creates a Prisoner’s Dilemma where Consumers feel forced into the high-pain $S_H$ strategy to avoid the $M$ (Mayonnaise) penalty, even if they would prefer $S_M$.
Nash Equilibria Analysis
This analysis treats the Spicy Food Semiotic Field as a Signaling Game with asymmetric information, where the Consumer’s “Type” (Tough/Authentic vs. Weak/Poser) is private information, and the Observer must infer this type through the “costly signal” of capsaicin consumption.
Part 1: Game Structure Analysis
1. Game Type
- Non-Cooperative: Players act in their own self-interest to maximize individual payoffs (Status vs. Social Accuracy).
- Sequential: The Consumer moves first (chooses heat level); the Observer moves second (chooses social response).
- Signaling Game (Bayesian): A subset of dynamic games with asymmetric information where one player (Sender) has information the other (Receiver) lacks.
2. Strategy Spaces
- Consumer ($S_C$):
- Extreme Heat: High-cost signal (Pain + Biological Honesty risk).
- Mild/Moderate: Low-cost signal (Risk of “Mayonnaise Proxy” penalty).
- Observer ($S_O$):
- Grant Status: Social validation, respect, or inclusion.
- Withhold Status: Mockery, dismissal, or “Mayonnaise” labeling.
- Constraints: Biological Honesty acts as a “leaky” constraint. A Consumer choosing “Extreme Heat” cannot fully control involuntary physiological signals (sweat, tears), which may reveal them as a “Weak” type attempting to bluff.
3. Payoffs
- Consumer Payoff ($U_C$): $Social Capital + Endorphin Loop - (Physical Pain + Biological Honesty Failure)$.
- Observer Payoff ($U_O$): $Accuracy of Interpretation - Social Effort$. The Observer gains utility by correctly identifying “Authentic” types and loses utility by being “duped” by a “Poser” or by appearing elitist/wrong.
4. Key Features
- Information Asymmetry: The Consumer knows their TRPV1 receptor density (genetic tolerance); the Observer does not.
- The Mayonnaise Proxy: A negative constant applied to the “Mild” strategy in specific subcultures, increasing the pressure to signal.
- Costly Signaling: For the signal to be “honest,” the pain of Extreme Heat must be higher for a “Weak” type than for a “Tough” type, making it harder for the Weak type to mimic the Tough type.
Part 2: Payoff Matrix (Simplified)
| Consumer Type | Strategy | Observer: Grant Status | Observer: Withhold Status |
|---|---|---|---|
| Tough (T) | Extreme Heat | (High, High) | (Med, Low) |
| Tough (T) | Mild | (Low, Low) | (Negative, Med) |
| Weak (W) | Extreme Heat | (Med*, Negative) | (Negative**, High) |
| Weak (W) | Mild | (Low, Low) | (Negative***, Med) |
*Med for Consumer due to high pain; Negative for Observer for being duped. **Negative for Consumer due to pain + mockery; High for Observer for “catching” a poser. ***The “Mayonnaise Proxy” penalty.
Part 3: Nash Equilibrium Analysis
In signaling games, we look for Perfect Bayesian Equilibria, categorized into “Separating” and “Pooling” equilibria.
Equilibrium 1: The “Authentic” Separating Equilibrium
- Strategy Profile:
- Tough Consumer: Consumes Extreme Heat.
- Weak Consumer: Consumes Mild.
- Observer: Grants Status if Heat is consumed; Withholds Status if Mild is consumed.
- Why it’s a Nash Equilibrium:
- The Tough type gains status and endorphins; they have no incentive to switch to Mild (losing status).
- The Weak type avoids the “Biological Honesty” failure. The pain of the heat + the risk of being caught “faking it” (leading to mockery) is worse than the “Mayonnaise Proxy” penalty of choosing Mild.
- The Observer correctly identifies both types, maximizing their accuracy payoff.
- Classification: Pure Strategy Separating Equilibrium.
- Stability/Likelihood: High in culinary subcultures where “authenticity” is valued over raw performance. It is stable because the “cost” of the signal (pain) is high enough to prevent the Weak type from mimicking the Tough type.
Equilibrium 2: The “Hot-Wing Industrial Complex” Pooling Equilibrium
- Strategy Profile:
- Both Types (Tough & Weak): Consume Extreme Heat.
- Observer: Grants Status to everyone (or grants status with a fixed probability).
- Why it’s a Nash Equilibrium:
- If the “Mayonnaise Proxy” penalty is sufficiently high (social death for eating mild food), the Weak type will endure extreme pain to avoid the penalty.
- The Tough type continues to eat heat because they enjoy the endorphins and the status.
- The Observer cannot distinguish between types and defaults to granting status to avoid the social cost of “gatekeeping” or because the spectacle itself is the value.
- Classification: Pure Strategy Pooling Equilibrium.
- Stability/Likelihood: High in “Challenge” cultures (e.g., YouTube, frat houses). It is stable as long as the social stigma of “Mild” is greater than the physical agony of “Extreme.”
Equilibrium 3: The “Skeptical” Mixed Strategy Equilibrium
- Strategy Profile:
- Weak Consumer: Randomizes between Heat and Mild.
- Observer: Randomizes between Granting and Withholding Status when they see Heat.
- Why it’s a Nash Equilibrium: If the Observer becomes too cynical (assuming everyone is a poser), they withhold status. This causes some Weak types to stop trying. This reaches a point where the Observer is unsure if the person sweating is “Tough” or just a “Weak” person trying hard, leading to a probabilistic response.
- Classification: Mixed Strategy Equilibrium.
- Stability/Likelihood: Common in “Hipster” or “Elite” food circles where “trying too hard” is mocked. It is less stable due to high coordination costs.
Part 4: Discussion of Multiple Equilibria
- Most Likely to Occur: The Pooling Equilibrium is increasingly dominant in the digital age. The “Hot-Wing Industrial Complex” has raised the “Mayonnaise Proxy” penalty so high that the “Mild” strategy is effectively removed from the strategy space for anyone seeking social relevance.
- Coordination Problems: The “Biological Honesty” constraint creates a coordination problem. If a Weak type attempts the Pooling Equilibrium (Extreme Heat) but their body betrays them (vomiting/fainting), the Observer may unilaterally switch to “Withhold Status,” collapsing the Pooling Equilibrium into a “Failed Separating” state.
- Pareto Dominance:
- The Separating Equilibrium is Pareto Superior to the Pooling Equilibrium. In the Separating Equilibrium, the Weak type does not have to suffer physical pain, and the Observer gets accurate information.
- The Pooling Equilibrium is Pareto Inferior because it forces the Weak type to incur high physical costs (Pain) for a diluted social reward (since the Observer knows the signal might be a bluff). However, the “Mayonnaise Proxy” creates a “Prisoner’s Dilemma” effect where players are forced into the inferior Pooling outcome to avoid the worst-case social stigma.
Dominant Strategies Analysis
This analysis explores the Spicy Food Semiotic Field through the lens of formal game theory, specifically as a Signaling Game with asymmetric information and costly signals.
Part 1: Game Structure Analysis
- Game Type:
- Non-cooperative: Players act in their own self-interest (Consumer seeks status/pleasure; Observer seeks to gatekeep/validate social norms).
- Sequential: The Consumer moves first (chooses heat level); the Observer moves second (interprets and reacts).
- Signaling Game: A sub-type of Bayesian games where one player (Consumer) has private information (their true toughness/tolerance) and sends a signal (consuming heat) to an uninformed player (Observer).
- Information Structure:
- Asymmetric/Imperfect Information: The Consumer knows their internal pain threshold and “authenticity.” The Observer only sees the external “Biological Honesty” (sweat, tears) and the Scoville level.
- Biological Honesty Constraint: This acts as a “Hard Signal.” Unlike verbal claims, physiological reactions are difficult to bluff, reducing the “cheap talk” element of the game.
- Players:
- Consumer (C): The Sender.
- Observer (O): The Receiver.
- Asymmetries:
- Payoff Asymmetry: The Consumer risks physical pain and biological distress. The Observer risks only “social miscalculation” (granting status to a “poser” or mocking a truly “tough” individual).
Part 2: Strategy Spaces
- Consumer Strategy Space ($S_C$):
- $S_{High}$ (Extreme Heat): High physical cost, high signal strength.
- $S_{Low}$ (Mild/Moderate): Low physical cost, low signal strength (risks the “Mayonnaise Proxy” penalty).
- Constraint: The Consumer cannot choose to not sweat if they choose $S_{High}$ and lack the genetic tolerance (Biological Honesty).
- Observer Strategy Space ($S_O$):
- $A_{Grant}$ (Respect/Validation): Bestows social capital.
- $A_{Withhold}$ (Mockery/Dismissal): Imposes social stigma or ignores the signal.
Part 3: Characterize Payoffs
The payoffs are non-transferable and involve both biological and social variables.
- Consumer Payoff ($U_C$):
$U_C = (Social Capital + Endorphin Loop) - (Physical Pain + Mayonnaise Proxy Penalty)$
- Note: The “Mayonnaise Proxy” is a negative payoff applied only if $S_{Low}$ is chosen in a high-stakes social subculture.
- Observer Payoff ($U_O$):
$U_O = (Social Order Maintenance) - (Credibility Cost)$
- The Observer gains utility by correctly identifying “Toughness” and loses utility if they are “fooled” by a performer or if they appear “weak” by association.
Payoff Matrix (Simplified)
| Consumer \ Observer | Grant Status ($A_G$) | Withhold Status ($A_W$) | | :— | :— | :— | | Extreme Heat ($S_H$) | $(SC + E) - P, \text{Validation}$ | $(E - P), \text{Mockery/Skepticism}$ | | Mild/Moderate ($S_M$) | $0, \text{Neutral}$ | $-MP, \text{Dismissal}$ |
(Where $SC$=Social Capital, $E$=Endorphins, $P$=Pain, $MP$=Mayonnaise Proxy)
Part 4: Key Features
- The Handicap Principle: Extreme heat consumption is a “Handicap” signal. Because it is genuinely painful ($P$), only those who value the status ($SC$) or have high tolerance (lower $P$) can afford to send the signal.
- The Endorphin Loop (Private Payoff): This creates a “floor” for the Consumer’s payoff. Even if the Observer withholds status, the Consumer may still receive a biological reward ($E$), making the game not strictly zero-sum.
- Timing: The “Biological Honesty” (sweating/flushing) occurs during the move, providing the Observer with real-time data to decide between $A_G$ and $A_W$.
Part 5: Dominant Strategy Analysis
1. Strictly Dominant Strategies
- Consumer: None.
- If the pain ($P$) is greater than the sum of social capital ($SC$) and endorphins ($E$), $S_H$ is not dominant.
- If the social subculture does not punish mildness (no $MP$), $S_M$ is not dominant.
- Observer: None.
- The Observer’s best response depends entirely on the Consumer’s “Biological Honesty.” If the Consumer looks like they are dying, the Observer may choose $A_W$ (Mockery) under the Performance Hypothesis.
2. Weakly Dominant Strategies
- Consumer: $S_H$ (Extreme Heat) becomes weakly dominant in “Chili-head” or “Macho” subcultures.
- In these environments, the “Mayonnaise Proxy” ($MP$) is a guaranteed negative payoff for $S_M$. Choosing $S_H$ at least offers a probability of $SC$ and a guaranteed $E$, even if $P$ is high.
- Observer: $A_W$ (Withhold/Dismiss) can be weakly dominant for an Observer who values “Gatekeeping” above all else. By withholding status, they protect their own social standing from being “cheapened” by over-validating others.
3. Dominated Strategies
- Consumer: $S_M$ (Mild) is a dominated strategy in high-status competitive environments (e.g., a “Hot Wing Challenge”).
- Choosing $S_M$ results in a payoff of $-MP$ (social death) with zero chance of $SC$ or $E$. Even a painful $S_H$ has a higher expected utility.
4. Iteratively Eliminated Strategies
- If we assume a subculture where the “Mayonnaise Proxy” is extremely high, we eliminate $S_M$ for the Consumer.
- Knowing the Consumer must choose $S_H$ to survive socially, the Observer then eliminates $A_G$ (Grant Status) for anyone who shows too much biological distress, as they are interpreted as “Performers” rather than “Naturally Tough.”
- This leads to a Separating Equilibrium where only those with high genetic tolerance (low $P$) receive status.
Strategic Implications
- The “Toughness” Trap: In subcultures where $S_M$ is dominated (the Mayonnaise Proxy is active), Consumers are forced into a “Race to the Bottom” (or “Race to the Top” of the Scoville scale). This leads to the Industrialization of Suffering, where heat levels must continually increase to maintain signal strength.
- The Authenticity Paradox: Because “Biological Honesty” is a constraint, the most successful strategy for a Consumer is to actually possess high tolerance. If the signal (heat) is too easy to mimic, the Observer will switch to $A_W$ to avoid being a “sucker.”
- Social Cohesion vs. Individual Status: The “Ritual of Suffering Together” suggests a shift from a non-cooperative signaling game to a Coordination Game. If everyone chooses $S_H$, the group achieves a “Pooling Equilibrium” where the shared pain creates a collective payoff, neutralizing the individual mockery ($A_W$).
- The Blackout Effect: Strategically, the “Sophistication Paradox” shows that as Consumers chase the $SC$ of $S_H$, they sacrifice the utility of “Flavor.” The game theory suggests that in the Spicy Food Semiotic Field, social capital is often valued more than the primary utility of the food itself.
Pareto Optimality Analysis
This analysis explores the Spicy Food Semiotic Field through the lens of game theory, specifically as a Signaling Game with elements of Costly Signaling Theory.
Part 1: Game Structure Analysis
1. Identify the Game Structure
- Type: Non-cooperative. While players exist in a shared social space, their objectives are often at odds (the Consumer wants status for the least effort/pain; the Observer wants to gatekeep status to maintain its value).
- Form: Sequential with Imperfect Information. The Consumer moves first (chooses heat level). The Observer moves second (interprets the signal). The “Imperfect Information” stems from the Observer not knowing the Consumer’s true biological tolerance (their “Type”).
- Game Length: Can be One-shot (a “Hot Wing Challenge” at a bar) or Repeated (building a reputation within a subculture).
- Asymmetries:
- Information Asymmetry: The Consumer knows their own pain threshold; the Observer only sees the “Biological Honesty” (sweat, tears).
- Payoff Asymmetry: The Consumer risks physical pain ($P$); the Observer only risks social misjudgment.
2. Define Strategy Spaces
- Consumer ($C$):
- Extreme Heat: High Pain, High Signal. (Discrete choice, though Scoville levels offer a continuous scale).
- Mild/Moderate: Low Pain, Low Signal.
- Observer ($O$):
- Grant Status: Respect, validation of “Toughness” or “Authenticity.”
- Withhold Status: Mockery (The Mayonnaise Proxy), dismissal of the performance as “fake” or “trying too hard.”
- Constraints: Biological Honesty. A Consumer cannot choose “Extreme Heat” and “No Physiological Reaction” simultaneously unless they have a specific genetic “Type.” The body’s involuntary response acts as a “leakage” that constrains the ability to bluff.
3. Characterize Payoffs
- Consumer Objectives: Maximize Social Capital ($S$) and Endorphin Loop ($E$), while minimizing Physical Pain ($P$) and avoiding the Mayonnaise Proxy ($M$).
- Payoff Function: $U_c = S + E - P - M$
- Observer Objectives: Maximize the “Purity” of the subculture. They gain utility by correctly identifying “Authentic” consumers and lose utility by granting status to “Posers” or “Weak” types.
- Transferability: Payoffs are non-transferable. The Consumer cannot “give” their endorphins to the Observer; the Observer cannot “give” their social standing to the Consumer without losing some of their own gatekeeping authority.
4. Key Features
- Signaling: The heat is a “Costly Signal.” Because it hurts, it is difficult for “Weak Types” to mimic “Tough Types.”
- The Endorphin Loop: This acts as a Private Payoff. Even if the Observer withholds status, the Consumer may still receive a biological reward, creating a “floor” for the payoff.
- The Mayonnaise Proxy: A negative externality for choosing the “Safe” strategy. In certain subcultures, choosing Mild is not a neutral move ($0$); it is a negative move ($-M$).
Part 2: Pareto Optimality Analysis
To analyze Pareto outcomes, we represent the interaction in a simplified payoff matrix. Assume: $S=10, E=2, P=5, M=8$.
| Consumer \ Observer | Grant Status | Withhold Status |
|---|---|---|
| Extreme Heat | (7, 5) [Status + Endorphins - Pain] | (-11, 2) [Mockery + Endorphins - Pain] |
| Mild/Moderate | (2, 0) [Base Acceptance] | (-8, 5) [Mayonnaise Proxy / Gatekeeper Win] |
1. Identification of Pareto Optimal Outcomes
An outcome is Pareto optimal if no player can be made better off without making the other worse off.
- (Extreme Heat, Grant Status) [7, 5]: Pareto Optimal. The Consumer gets high status and endorphins; the Observer gets the satisfaction of witnessing a successful performance.
- (Mild/Moderate, Withhold Status) [-8, 5]: Pareto Optimal. While the Consumer is miserable (Mayonnaise Proxy), the Observer is at their maximum utility for successfully gatekeeping and reinforcing subculture boundaries.
- (Mild/Moderate, Grant Status) [2, 0]: Pareto Optimal. The Consumer avoids pain and mockery; the Observer is neutral.
2. Comparison to Nash Equilibria
In a “Toughness” subculture (where $M$ is high):
- Nash Equilibrium: (Extreme Heat, Grant Status). If the Consumer knows the Observer will mock mildness, they are forced into Extreme Heat. If the Consumer eats Extreme Heat, the Observer’s best response (to avoid looking like a hater or to enjoy the spectacle) is to Grant Status.
- Alignment: In this specific subculture, the Nash Equilibrium is also a Pareto Optimal outcome. However, it is “Pain-Inefficient” for the Consumer.
3. Pareto Improvements
A Pareto improvement occurs when we can make one person better off without hurting the other.
- From (Extreme Heat, Withhold) to (Extreme Heat, Grant): If the Consumer is already in pain, the Observer granting status makes the Consumer better off ($7$ vs $-11$) and the Observer better off ($5$ vs $2$).
- The “Authenticity” Trap: There is often no Pareto improvement from (Extreme Heat, Grant) to (Mild, Grant) because while the Consumer’s pain decreases, the Observer’s utility decreases because the “signal” has been devalued. The Observer loses the “Social Poison” utility of judging a difficult feat.
4. Efficiency vs. Equilibrium Trade-offs
- The Sophistication Paradox: The Nash Equilibrium of “Extreme Heat” is often socially inefficient. Both players spend resources (Consumer spends physical health/comfort; Observer spends cognitive energy policing) to reach a state that could be achieved through “Mild/Moderate” consumption if the “Mayonnaise Proxy” were removed.
- Coordination Failure: The “Mayonnaise Proxy” creates a Stag Hunt dynamic. If everyone agreed to stop mocking mild food, everyone could enjoy flavor without pain. However, the first person to “defect” and eat spicy food gains a massive status boost, dragging the group back into the “Spicy Food Semiotic Field.”
Conclusion: Opportunities for Cooperation
To reach a Pareto improvement that reduces total pain (The “Sensory Peace” outcome), players must engage in Pre-play Communication:
- Redefining the Signal: Shift the “Authenticity” metric from Heat (Pain) to Complexity (Flavor). This moves the game from a “Toughness Hypothesis” (Zero-sum) to a “Connoisseurship Game” (Cooperative).
- De-weaponizing the Mayonnaise Proxy: By removing the negative payoff for Mild strategies, the Consumer is no longer “forced” into the Extreme strategy by the threat of mockery, allowing for a Nash Equilibrium at (Mild, Grant) which is more physically efficient.
Strategic Recommendations
This analysis explores the Spicy Food Semiotic Field through the lens of game theory, treating the consumption of capsaicin as a signaling game where biological distress functions as “Proof of Work.”
Part 1: Game Structure Analysis
1. Identify the Game Structure
- Type: Non-cooperative Signaling Game.
- Timing: Sequential. The Consumer moves first (chooses heat level); the Observer moves second (interprets the signal and assigns status).
- Information: Imperfect and Asymmetric. The Consumer knows their true tolerance and internal pain level; the Observer only sees the outward “Biological Honesty” (sweat, flush) and the heat level of the food.
- Repetition: Can be one-shot (a “Hot Wing Challenge”) or repeated (building a reputation in a subculture).
2. Define Strategy Spaces
- Consumer ($C$):
- Consume Extreme Heat ($S_H$): High physical cost, high potential signal.
- Consume Mild/Moderate ($S_M$): Low physical cost, low signal/potential social penalty.
- Observer ($O$):
- Grant Status ($A_G$): Validation of toughness or authenticity.
- Withhold Status ($A_W$): Mockery, dismissal, or labeling with the “Mayonnaise Proxy.”
- Constraints: “Biological Honesty” acts as a signal-jamming constraint. A Consumer cannot easily suppress a flush (involuntary), making it a “costly signal” that is difficult to fake.
3. Characterize Payoffs
- Consumer Payoff: $U_C = (Endorphin Loop + Social Capital) - (Physical Pain + Social Penalty)$.
- The “Mayonnaise Proxy” is a heavy negative weight applied to $S_M$ in specific subcultures.
- Observer Payoff: $U_O = (Accuracy of Judgment) - (Social Effort)$.
- The Observer gains utility by correctly identifying a “tough” individual (Toughness Hypothesis) or an “authentic” one (Performance Hypothesis) and loses utility if they are “fooled” by a poser.
4. Key Features
- Signaling: The heat level is the signal. The physiological reaction (sweat/tears) is the “verification” of the signal’s honesty.
- The Sophistication Paradox: A sub-game where choosing too much heat results in a negative payoff for “Authenticity” because it suggests the Consumer can no longer taste the food.
Part 2: Strategic Recommendations
For the Consumer
- Optimal Strategy: The “Controlled Burn” (High Heat + Visible Resilience).
- Play $S_H$ (Extreme Heat) only when the “Endorphin Loop” (private payoff) is high enough to offset the “Physical Pain.” To maximize “Social Capital,” you must exhibit “Biological Honesty” (sweating) but maintain “Stoic Performance” (not complaining).
- Contingent Strategies:
- If the Observer is an “Authenticity Cop”: Choose “Thai/Indian Spicy” rather than “Extract-based Heat.” This signals cultural capital rather than mere masochism.
- If the Observer uses the “Mayonnaise Proxy”: Avoid $S_M$ at all costs to prevent a permanent status downgrade.
- Risk Assessment:
- The “Try-Hard” Trap: If the signal is too high (e.g., ordering the “Mega-Death Sauce”) but the physiological reaction is catastrophic (vomiting), the Observer switches from $A_G$ (Respect) to $A_W$ (Mockery).
- Coordination Opportunities: Engage in “Rituals of Suffering Together.” By consuming heat with the Observer, you transform a non-cooperative signaling game into a cooperative bonding exercise.
- Information Considerations: Use “Biological Honesty” to your advantage. Do not wipe away the sweat immediately; let the Observer see the “Proof of Work” to validate the signal’s cost.
For the Observer
- Optimal Strategy: “Skeptical Validation.”
- Withhold status ($A_W$) initially to force the Consumer to increase the signal’s cost. Grant status ($A_G$) only when the “Biological Honesty” (involuntary reaction) confirms the Consumer is not “bluffing” with a high genetic tolerance.
- Contingent Strategies:
- Against a “Poser”: If the Consumer orders high heat but shows no reaction, invoke the “Sophistication Paradox”—accuse them of destroying their palate and withhold status.
- Risk Assessment:
- False Negative: Mocking someone who is genuinely enduring extreme pain can lead to social blowback for the Observer (appearing elitist or cruel).
- Coordination Opportunities: Use the “Mayonnaise Proxy” as a gatekeeping tool to maintain the “purity” and high-entry-cost of the subculture.
- Information Considerations: Look for “Micro-expressions” of distress. The gap between the Consumer’s “Performance” (words) and “Biology” (tears) is where the most accurate information resides.
Part 3: Overall Strategic Insights
- The Costly Signaling Equilibrium: Social status in the spicy food field is only granted when the cost (pain) is visible. If a Consumer has a naturally high tolerance (low cost), their signal is “cheaper” and may actually result in less status than someone who struggles visibly but persists.
- The “Mayonnaise” Deterrent: The “Mayonnaise Proxy” creates a “Prisoner’s Dilemma” where both players might prefer a mild meal, but both choose spicy to avoid the social stigma of being “bland.”
Part 4: Potential Pitfalls
- Over-Signaling: Choosing a heat level that triggers a “Biological Honesty” response so severe it breaks the “Performance” (e.g., crying or needing medical attention). This results in a total loss of Social Capital.
- Misreading the Subculture: Using “Toughness” signals in a “Sophistication” environment. In a high-end culinary setting, sweating over a ghost pepper is seen as “low-status” rather than “tough.”
Part 5: Implementation Guidance
- For Consumers: If you must play the game, build “Desensitization Capital” privately. This lowers your internal “Cost of Pain” while allowing you to still trigger “Biological Honesty” signals for the Observer.
- For Observers: To be a high-status judge, focus on the “Sensory Semiotic Triad.” Contrast the Consumer’s reaction to Spicy (public) with their reaction to Bitter (private) to determine if they are a “Performer” or a true “Chili-head.”
Game Theory Analysis Summary
GameAnalysis(game_type=Signaling Game (Asymmetric Information) with elements of a Status/Prestige Game, players=[The Consumer (The Signaler), The Social Group/Observer (The Receiver)], strategies={The Consumer=[Biological Honesty (Low/Moderate Heat), The Performance (High Heat), Authenticity Policing], The Social Group/Observer=[Validation, Derision (The Mayonnaise Proxy), Skepticism]}, payoff_matrix=Consumer Payoff: Social Capital + Cultural Authenticity - (Physical Pain + Sensory Blackout). Observer Payoff: Social Cohesion + Entertainment Value - Information Uncertainty. Sophistication Paradox: Status gain vs. sensory loss., nash_equilibria=[The ‘Toughness’ Equilibrium: All Consumers choose High Heat to avoid the ‘Mayonnaise’ label in group settings., The ‘Hot-Wing Industrial’ Equilibrium: Media creators provide extreme pain and audiences provide views/validation in an escalation cycle.], dominant_strategies={The Consumer=High Heat (in competitive social environments), The Social Group/Observer=Over-interpretation}, pareto_optimal_outcomes=[Biological Honesty: Consumer eats at true biological comfort level and Observer grants status based on culinary knowledge., Total Utility: Maximizes Consumer’s physical enjoyment and Observer’s social interaction without physical suffering or interpretive overfitting.], recommendations={The Consumer=Recognize the Sophistication Paradox and decouple self-worth from TRPV1 receptor density., The Social Group (Observer)=Avoid the Mayonnaise Proxy and acknowledge that ‘quiet’ tastes are equally complex., The ‘Chili-Head’ Community=Shift the game from Endurance (Pain) to Discernment (Flavor) to move toward a more Pareto-efficient experience.})
Analysis completed in 132s Finished: 2026-02-20 20:10:52
Socratic Dialogue Transcript
Initial Question: If the capsaicin response is a ‘biological absolute’ of pain, does its transition into a social ‘semiotic field’ represent a corruption of its honesty, or is the very concept of ‘biological honesty’ a social construct used to validate performative suffering?
Domain Constraints: Biology, Sociology, Semiotics, Cultural Theory
Started: 2026-02-20 20:10:37
Exchange 1
Question: If the capsaicin response is a ‘biological absolute’ of pain, does its transition into a social ‘semiotic field’ represent a corruption of its honesty, or is the very concept of ‘biological honesty’ a social construct used to validate performative suffering?
Response: To address this question, we must first dismantle the components of the “capsaicin response” and examine how biology and culture intersect at the site of the nerve ending.
1. Defining the “Biological Absolute”
From a biological perspective, capsaicin is a chemical irritant that binds to the TRPV1 (transient receptor potential vanilloid 1) ion channel. These receptors are primarily responsible for detecting and regulating body temperature, as well as providing a sensation of scalding heat and physical abrasion.
When we call this a “biological absolute,” we are referring to the nociceptive signal: the chemical-to-electrical transduction that tells the brain “this is hot/painful.” However, biology is rarely “absolute” in its interpretation. The brain’s processing of that signal—the transition from nociception to the experience of pain—is subject to neuroplasticity and habituation. Therefore, while the chemical binding is a biological fact, the “pain” is already a subjective interpretation by the nervous system before it even reaches the level of social discourse.
2. The Semiotic Field: Corruption or Evolution?
The transition of capsaicin into a “semiotic field” occurs when the sensation of burning is detached from its evolutionary purpose (warning of tissue damage) and reattached to a new set of meanings (culinary sophistication, masculinity, or cultural heritage).
Does this represent a corruption of honesty? If we define “honesty” as the alignment between a signal and its evolutionary intent, then yes, enjoying a habanero is a “lie.” The body is signaling a life-threatening thermal event that is not actually occurring. We are “hacking” the nervous system to produce an endorphin rush.
However, in semiotics, a sign is rarely “honest” in a vacuum; it is functional. By moving the capsaicin response into a semiotic field, we are not necessarily corrupting it, but rather re-coding it. The “burn” becomes a signifier for “authenticity” or “intensity.” The “honesty” is not lost; it is redirected from a biological warning to a social signal.
3. The Social Construction of “Biological Honesty”
The most provocative part of your question is whether “biological honesty” is a construct used to validate performative suffering.
In many cultures, pain is viewed as the “ultimate truth” because it is perceived as being beyond the reach of the will. We see this in:
- The “Hot Ones” phenomenon: The celebrity interview becomes “honest” only when the subject is in physical distress, under the assumption that the “biological absolute” of pain bypasses their PR-trained persona.
- Rites of passage: Using spicy peppers or physical trials to prove “manhood” or “strength.”
Here, “biological honesty” is indeed a social construct. We imbue the physiological response with a moral weight. We decide that the involuntary sweating and gasping caused by capsaicin are “more real” than the words the person is speaking. This validates the performance of suffering as a metric of character.
4. Synthesis and Implications
I would argue that the concept of “biological honesty” is a strategic essentialism. We pretend the body cannot lie so that we have a fixed point of reference in an increasingly performative world.
However, the capsaicin response is uniquely suited for this because it is a “safe” pain. Unlike a broken bone or a disease, capsaicin causes no actual tissue damage (in moderate amounts). This makes it the perfect medium for “performative honesty”: it provides all the visual and physiological markers of a biological crisis without the actual biological cost.
Initial Conclusion: The transition of the capsaicin response into a semiotic field is not a corruption, but a domestication of the absolute. We take a raw biological signal and turn it into a tool for social communication. The “honesty” we attribute to it is a cultural layer we apply to make sense of our desire to suffer for pleasure or status.
A question for further exploration: If we acknowledge that we use “biological honesty” to validate performance, does that mean all shared cultural experiences of pain (from spicy food to marathons) are fundamentally theatrical, or is there a point where the biology becomes so overwhelming that the semiotic field collapses?
Next Question: If the nervous system’s transition from a chemical signal to the experience of pain is already a subjective interpretation, does the “biological absolute” function as a ground for truth, or is it merely the first layer of semiotic mediation in which the body “interprets” its environment?
Exchange 2
Question: If the nervous system’s transition from a chemical signal to the experience of pain is already a subjective interpretation, does the “biological absolute” function as a ground for truth, or is it merely the first layer of semiotic mediation in which the body “interprets” its environment?
Response: To address whether shared cultural experiences of pain are fundamentally theatrical or if they reach a point of semiotic collapse, we must examine the boundary between the symbolic order (where we assign meaning) and the real (the raw, unmediated biological drive).
1. The “Theatricality” of Structured Suffering
In sociology, specifically through the lens of Erving Goffman’s “Dramaturgy,” we can argue that most cultural experiences of pain are indeed theatrical—but this does not mean they are “fake.” Instead, they are framed.
A marathon runner and a person fleeing a predator may experience identical physiological markers: elevated cortisol, lactic acid buildup, and extreme cardiovascular stress. However, the runner’s pain is “theatrical” because it exists within a specific semiotic frame (the race, the medal, the personal best). The frame provides a “script” for the pain. Without this script, the pain is merely trauma; with it, the pain is “endurance.”
In this sense, the “theatricality” is the very thing that allows the pain to be shared. By turning the biological signal into a performance, we make it legible to others. We are not just feeling; we are communicating.
2. The Threshold of Semiotic Collapse
However, your question asks if there is a point where the biology becomes so overwhelming that this semiotic field collapses. I would argue that this point exists and is defined by the limit of language.
In her seminal work The Body in Pain, Elaine Scarry argues that physical pain does not simply resist language but actively destroys it. Intense pain brings about a “reversion to a state anterior to language,” reducing the sufferer to moans, cries, and non-verbal sounds.
The Collapse occurs when:
- Cognitive Resource Exhaustion: The prefrontal cortex (responsible for meaning-making, social filtering, and “performance”) is bypassed by the autonomic nervous system.
- The Loss of the “Other”: In a theatrical experience of pain, there is always an implied audience (even if it is just the self-observing the self). In total semiotic collapse, the “audience” vanishes. There is no “I am suffering for a reason”; there is only “Suffering is.”
At this point, the capsaicin is no longer a “signifier of authenticity” or a “challenge.” It becomes a totalizing biological event that consumes the subject’s entire reality. The “Hot Ones” interview ceases to be an interview; the marathon runner collapses into a seizure. The semiotic field hasn’t just been “corrupted”—it has been annihilated.
3. The “Aesthetic of the Edge”
The most interesting cultural phenomenon is not the collapse itself, but our collective fascination with the approach to that collapse.
We engage in “safe” versions of this collapse—what we might call controlled de-semiotization. We want to see the celebrity on “Hot Ones” lose their composure because we want to see the “mask” (the semiotic persona) fail. We are looking for the moment where the “biological absolute” breaks through the “cultural performance.”
This is a pursuit of the Sublime, as defined in aesthetic theory (Burke/Kant): the experience of something so vast or powerful that it threatens to overwhelm our capacity to process it, yet we find pleasure in it because we experience it from a position of relative safety.
4. Synthesis: The Paradox of Shared Pain
If the pain is shared and cultural, it is, by definition, semiotic. The moment it becomes “real” (the collapse), it becomes unshareable.
True biological collapse is profoundly isolating. You cannot “share” a grand mal seizure or the peak of a torture session in a way that maintains a cultural narrative; those events exist outside the bounds of social exchange. Therefore:
- Shared cultural pain is a performance of “flirting with the collapse.”
- Theatricality is the bridge that allows us to experience the “biological absolute” without being destroyed by it.
Revised Understanding: We do not use “biological honesty” to validate performance; rather, we use performance to domesticate the biological absolute. We create rituals (spicy food, marathons, rites of passage) to touch the “Real” of our biology without falling into the abyss of total semiotic dissolution.
A question for further exploration: If the “truth” of the body is only found at the point where language and meaning collapse, does this imply that our entire “civilized” existence is a flight from biological reality? If we only feel “honest” when we are in a state of semiotic breakdown, what does that say about the “honesty” of our thoughts, emotions, and social structures?
Next Question: If the “collapse” into non-verbal cries and autonomic responses is considered a state of “semiotic dissolution,” how do we account for the fact that these “primitive” signals are often the most universally legible and biologically efficient forms of communication we possess?
Exchange 3
Question: If the “collapse” into non-verbal cries and autonomic responses is considered a state of “semiotic dissolution,” how do we account for the fact that these “primitive” signals are often the most universally legible and biologically efficient forms of communication we possess?
Response: To resolve the paradox you’ve identified—that the “collapse” of language results in the most “efficient” communication—we must distinguish between different modes of semiotics.
The “dissolution” I referenced previously refers specifically to the Symbolic Order (the realm of language, convention, and arbitrary signs). However, as you rightly point out, when the symbolic order fails, a more ancient and robust system takes over.
To account for the universal legibility of these “primitive” signals, we can look through the following lenses:
1. From Symbol to Index (The Peircean Shift)
In the semiotics of C.S. Peirce, there is a crucial distinction between a Symbol and an Index:
- Symbols (like the word “pain”) are arbitrary and require a cultural “key” to decode. They are fragile and easily lost in the “collapse.”
- Indices (like a scream, a dilated pupil, or a sudden sweat) have a direct, causal connection to their object. Smoke is an index of fire; a gasp is an index of sudden physiological shock.
The “efficiency” you mention arises because indices do not require “translation.” When the symbolic field dissolves, we are not left with nothing; we are left with the Indexical Absolute. The reason these signals are universally legible is that they are not “read” by the linguistic centers of the brain (the Broca’s area), but are “felt” by the primitive brain (the amygdala and the periaqueductal gray).
2. Biological Efficiency: The “Fast Path”
From a biological standpoint, the “collapse” into non-verbal cries is an evolutionary feature, not a bug. In neurology, we speak of the “Low Road” vs. the “High Road” of stimulus processing.
- The High Road (Symbolic): Sensory data $\rightarrow$ Thalamus $\rightarrow$ Sensory Cortex $\rightarrow$ Prefrontal Cortex $\rightarrow$ Response. This is slow, nuanced, and culturally dependent.
- The Low Road (Primitive): Sensory data $\rightarrow$ Thalamus $\rightarrow$ Amygdala $\rightarrow$ Autonomic Response. This is instantaneous.
The “legibility” of a cry is efficient because it bypasses the “theatrical” processing of the observer. If I see you crying out in pain, my mirror neurons fire an empathetic response before I have even consciously identified why you are crying. The communication is “efficient” because it is a direct nervous-system-to-nervous-system transmission, bypassing the “lag” of cultural interpretation.
3. The Monosemic vs. Polysemic
Language is polysemic—words have multiple meanings and are prone to deception, irony, and metaphor. A “primitive” signal is monosemic—it generally means one thing.
- A blush cannot be easily faked.
- A pupil dilation is an honest report of the autonomic nervous system.
We find these signals “legible” because they are unambiguous. In a world of complex social performances, the “collapse” into the primitive provides a “ground truth.” This is why, in sociology, we often view these involuntary responses as the “leakage” of the true self through the social mask.
4. The Paradox of “Legibility” without “Content”
Here is where we must be careful: Legibility is not the same as understanding. While a scream is “universally legible” as a sign of distress, it is “semiotically empty” regarding the specifics of that distress.
- A scream tells me that you are suffering.
- Language tells me why you are suffering (e.g., “My mortgage is underwater,” or “I have a sharp pain in my left gallbladder”).
The “collapse” is a trade-off: we gain certainty and speed (the biological efficiency), but we lose complexity and specificity (the cultural narrative). We know that the person on “Hot Ones” is in agony, but the agony itself has no “content” other than its own intensity.
Synthesis: The “Pre-Social” Foundation
We can account for the efficiency of these signals by recognizing them as the infrastructure upon which the “theatre” of culture is built. Sociologically, these primitive signals are the “fail-safes” of the human species. If our cultural symbols fail—if we encounter someone whose language we don’t speak or whose values we don’t share—the “collapse” into the primitive allows us to return to a baseline of shared biological reality.
Revised Understanding: The “collapse” is not a descent into chaos, but a regression to a more stable, lower-energy state of communication. We trade the “high-definition” but “unreliable” signal of language for the “low-definition” but “indisputable” signal of biology.
A question for further exploration: If these primitive, indexical signals are the most “honest” and “efficient” forms of communication, why did we evolve the “fragile” and “deceptive” symbolic order of language at all? If the “truth” of the body is so legible, what is the evolutionary or sociological advantage of the “mask” that language provides? Is the “theatricality” of culture a way to protect us from the overwhelming efficiency of the biological real?
Next Question: If the “Indexical Absolute” can be simulated—as in the case of a calculated sob, a forced laugh, or the trained suppression of pain—does this not suggest that the “primitive” signal is itself a site of potential deception, thereby collapsing the distinction between the “honest” biological report and the “theatrical” social mask?
Exchange 4
Question: If the “Indexical Absolute” can be simulated—as in the case of a calculated sob, a forced laugh, or the trained suppression of pain—does this not suggest that the “primitive” signal is itself a site of potential deception, thereby collapsing the distinction between the “honest” biological report and the “theatrical” social mask?
Response: Your question strikes at the heart of the “crisis of the real.” If the Indexical Absolute—the supposedly “honest” report of the body—can be hijacked by the “theatrical” social mask, then the boundary between biology and performance becomes porous. This suggests that the “primitive” is not a sanctuary of truth, but merely another stage for representation.
To address whether this collapses the distinction between the “honest” report and the “theatrical” mask, we must examine the mechanics of deception through the lenses of biology, semiotics, and sociology.
1. The Biological “Leak”: The Involuntary vs. The Voluntary
While we can simulate a sob or suppress a flinch, biology provides a “hardware” constraint that is difficult to override completely. In neurology, we distinguish between the pyramidal tract (voluntary movement) and the extrapyramidal tract (involuntary/emotional expression).
- The Duchenne Distinction: A classic example is the “Duchenne smile.” A true smile of joy involves the involuntary contraction of the orbicularis oculi (muscles around the eyes), which most people cannot contract at will. A “theatrical” smile uses only the zygomatic major (the mouth).
- Micro-expressions: Even the most trained “mask” (such as a poker player or a spy) often suffers from “leakage.” These are micro-expressions—involuntary facial movements lasting a fraction of a second—that betray the underlying emotional state before the conscious mind can suppress them.
Reasoning: The distinction does not “collapse” biologically; rather, it becomes a contest of bandwidth. The “honest” biological report operates on a faster, more subterranean frequency than the “theatrical” mask. Deception is possible, but it is biologically “expensive” and often imperfect.
2. The Semiotics of the “Pseudo-Index”
In Peircean terms, a calculated sob is what we might call a Pseudo-Index.
- A True Index is caused by its object (Pain $\rightarrow$ Cry).
- A Pseudo-Index is a Symbol designed to look like an Index (Intent to Deceive $\rightarrow$ Calculated Cry).
The “collapse” you suggest is actually a semiotic migration. When we simulate a primitive signal, we are taking a sign that should be monosemic (meaning only one thing) and making it polysemic (open to interpretation).
However, the “Indexical Absolute” remains a useful concept because the receiver of the signal still processes it as an index. If I see you cry, my amygdala reacts to the form of the cry before my prefrontal cortex questions its sincerity. The “theatrical” mask relies entirely on the “honest” report’s pre-existing reputation for truth. If the distinction truly collapsed—if we assumed all cries were fake—the signal would lose all social and evolutionary utility.
3. Sociology: The “Arms Race” of Authenticity
From a sociological perspective, the ability to simulate the primitive is an essential tool for social navigation. Erving Goffman’s Dramaturgy suggests that we are always “on stage.”
- The Social Mask as Protection: The “theatrical” suppression of pain is often a requirement for social status or tribal belonging (e.g., “boys don’t cry”).
- The Deception-Detection Arms Race: Because humans are “social animals” who rely on cooperation, we have evolved highly sophisticated “cheater detection” mechanisms. We are hyper-attuned to the “uncanny valley” of human emotion—that slight dissonance when a laugh sounds “forced” or a sob feels “performative.”
The distinction doesn’t collapse; it escalates. As our ability to “fake” the primitive improves, our ability to “detect” the fake must also improve. The “theatrical” and the “honest” exist in a dialectical tension.
4. The Implications of the “Calculated Sob”
If we accept that the primitive signal can be simulated, we must acknowledge a chilling implication: The “Real” becomes a style.
In a world of high-level social performance, “authenticity” itself becomes a theatrical trope. We see this in “raw” social media posts or “unfiltered” political speeches. These are calculated attempts to trigger the observer’s “Indexical Absolute” response. This is what Jean Baudrillard called Simulacra: a copy of a feeling that has no original “honest” version behind it.
Synthesis: The “Residual Real”
Does the distinction collapse? I would argue no, but it becomes probabilistic rather than absolute.
We can no longer say, “This signal is 100% honest because it is biological.” Instead, we must say, “This signal has a high probability of being honest because the biological cost of faking it (pupil dilation, micro-tremors, skin conductance) is too high for the conscious mind to sustain.”
The “theatrical” mask is a layer of Symbolic complexity laid over the Indexical foundation. The mask can mimic the foundation, but it cannot become the foundation. The “honest” report is the baseline; the “theatrical” mask is the modulation.
Revised Understanding: The “Indexical Absolute” is not a guarantee of truth, but a threshold of credibility. Deception does not collapse the distinction; it exploits the distinction. The “theatrical” mask only works because we—as biological observers—are hard-wired to believe the “honest” report.
A question for further exploration: If we live in a culture where the “theatrical” mask becomes so perfected that it is indistinguishable from the “honest” report (e.g., through deepfakes or advanced pharmacological emotional control), what happens to the concept of empathy? If empathy is a “nervous-system-to-nervous-system” resonance, can we empathize with a “calculated” signal? Or does empathy require the certainty of a shared biological vulnerability?
Next Question: If the “theatrical” mask can be internalized so thoroughly—through “method” techniques, trauma, or deep social conditioning—that it triggers the very involuntary “extrapyramidal” markers (such as the Duchenne smile or micro-expressions) used to define the “honest” report, does the distinction between the “biological foundation” and the “symbolic modulation” remain a difference of kind, or is “honesty” itself merely a performance that has successfully bypassed the subject’s own conscious awareness?
Exchange 5
Question: If the “theatrical” mask can be internalized so thoroughly—through “method” techniques, trauma, or deep social conditioning—that it triggers the very involuntary “extrapyramidal” markers (such as the Duchenne smile or micro-expressions) used to define the “honest” report, does the distinction between the “biological foundation” and the “symbolic modulation” remain a difference of kind, or is “honesty” itself merely a performance that has successfully bypassed the subject’s own conscious awareness?
Response: To address the question of empathy in the face of a “perfected” theatrical mask, we must pivot from the perspective of the sender (the one wearing the mask) to the receiver (the one witnessing it).
If the mask is so well-constructed that it triggers the extrapyramidal markers of the “honest” report, the receiver’s biological systems will respond reflexively. This leads us to a profound tension between biological resonance and ontological truth.
1. The Biological Reflex: Empathy as a “Hackable” System
Empathy is not merely a conscious moral choice; it is a neurological reflex rooted in the mirror neuron system. When we observe another person’s micro-expressions or hear the specific frequency of a sob, our brains simulate those states.
- The Mirror Reflex: If a “calculated” signal is biologically perfect, the observer’s mirror neurons will fire. The observer will feel the resonance of the other’s “pain” regardless of whether that pain has an internal referent in the sender.
- The Autonomic Bypass: Empathy often bypasses the prefrontal cortex (the seat of judgment) and hits the limbic system directly. In this sense, empathy is “blind.” It is a biological “key” that fits into a “lock.” If the key is a masterfully forged duplicate, the door opens just as surely as if it were the original.
Reasoning: Empathy does not require the reality of the other’s state; it only requires the perceptual conviction of it. Therefore, we can—and frequently do—empathize with “calculated” signals (in cinema, theater, or sophisticated political performance).
2. Sincerity vs. Authenticity: The Internalized Mask
You raised the possibility of the mask bypassing the subject’s own conscious awareness (e.g., through Method acting or deep conditioning). Here, we must distinguish between Sincerity and Authenticity.
- Sincerity: The alignment between what a person says/shows and what they believe they feel. A person who has internalized their mask is “sincere”—they are not “lying” because they have successfully deceived themselves.
- Authenticity: The alignment between the persona and the “biological foundation” or “original self.”
If a subject internalizes a mask so thoroughly that they trigger their own extrapyramidal markers, they are experiencing a simulated authenticity. The empathy we feel for them is “real” because their physiological state is “real,” even if the origin of that state is a theatrical construct. In this scenario, the distinction between the “honest” and the “theatrical” doesn’t just collapse for the observer; it collapses for the subject as well.
3. The “Empathy Economy” and the Erosion of Trust
If we move from the biological to the sociological, the “perfected mask” creates a crisis of Social Capital.
Empathy is a form of social glue that relies on a “shared biological vulnerability.” If we live in a culture where we suspect that every “honest” report is a “calculated” signal, we enter a state of empathic fatigue or cynical detachment.
- The Retroactive Withdrawal: Think of the feeling when you discover a “heartbreaking” viral video was staged. The empathy you felt was biologically real in the moment, but upon the revelation of the “mask,” you feel a sense of “empathic theft.”
- The Defense Mechanism: To protect ourselves from this “theft,” we may begin to raise our “Indexical Threshold.” We stop empathizing with the “Duchenne smile” and start looking for even deeper, harder-to-fake markers, or we require a history of consistent behavior (long-term data) before granting empathy.
4. Does Empathy Require Shared Vulnerability?
The most chilling implication of your question is whether empathy can survive the death of vulnerability.
True biological honesty is rooted in the fact that the body cannot help but betray itself. This “betrayal” is a form of vulnerability—it is the “Indexical Absolute” that says, “I am not in total control of my representation.”
If the “theatrical” mask becomes perfect, control is absolute. If I can choose exactly when to trigger a Duchenne smile or a micro-expression of grief, I am no longer vulnerable; I am a broadcaster of signals.
Revised Understanding: Empathy, in its purest form, is a resonance with the uncontrollable nature of the other. If we suspect a signal is “calculated,” our empathy shifts from affective resonance (feeling with them) to cognitive perspective-taking (thinking about them). We might still “understand” their plight, but the “nervous-system-to-nervous-system” bond is severed because the “vulnerability” is recognized as a “performance.”
Synthesis: The “Ghost” in the Mirror
Can we empathize with a “calculated” signal? Biologically, yes. Our neurons are easily fooled by a high-fidelity simulation. Sociologically and Existentially, no. Once the “calculation” is suspected, the nature of the connection changes from a communion to a transaction.
The “theatrical” mask doesn’t destroy empathy; it instrumentalizes it. It turns empathy into a resource to be harvested rather than a bridge to be crossed.
A question for further exploration: If we accept that “authenticity” is becoming a “style” (a set of high-fidelity theatrical markers), does this mean that the only way to be truly “honest” in the future is through failure? Is the only “honest” report the one that is clumsy, poorly performed, and “unconvincing”—the signal that fails to trigger the mirror neurons perfectly? Does “truth” now reside only in the “glitch”?
Synthesis
This synthesis integrates the biological, sociological, and semiotic threads of the dialogue to address the nature of “biological honesty” and the performative aspects of human suffering.
1. Summary of Key Insights
- The Nociceptive-Interpretive Gap: A fundamental distinction exists between nociception (the chemical-to-electrical transduction of the TRPV1 receptor) and pain (the subjective experience). Biology provides the signal, but the brain—influenced by neuroplasticity and habituation—provides the interpretation.
- Semiotic Decoupling: Humans possess the unique ability to detach a biological signal from its evolutionary purpose (warning of tissue damage) and reattach it to social signifiers (masculinity, culinary status, or religious devotion).
- The “Hackability” of Empathy: Empathy is a biological reflex driven by the mirror neuron system. Because this system responds to perceived signals rather than “ontological truth,” it can be “hacked” by a sufficiently perfected performance (the “theatrical mask”).
- Internalized Performance: When a social performance is internalized through deep conditioning or “method” techniques, it can trigger involuntary extrapyramidal markers. At this point, the distinction between a “real” feeling and a “performed” one becomes biologically indistinguishable.
2. Assumptions Challenged or Confirmed
- Challenged: The Purity of Biology. The assumption that biological signals are “honest” or “absolute” was challenged. The dialogue suggests that even at the level of the nerve ending, “honesty” is subject to the brain’s modulation.
- Challenged: Performance as Deception. The assumption that a “social mask” is inherently a lie was challenged. If the mask triggers the same physiological cascades as the “authentic” state, the mask becomes a new biological reality.
- Confirmed: The Evolutionary Basis of Pain. The dialogue confirmed that while the meaning of pain is social, the mechanism (TRPV1) remains a biological constant, providing the “raw material” for semiotic construction.
3. Contradictions and Tensions Revealed
- Biological Resonance vs. Ontological Truth: A profound tension exists where an observer can experience genuine biological empathy for a “fake” signal. This suggests that our social cohesion is built on a system (empathy) that cannot distinguish between a “masterfully forged key” and the “original key.”
- The Extrapyramidal Paradox: If “involuntary” markers (like the Duchenne smile or micro-expressions) can be triggered by a conscious decision to “internalize a mask,” the very definition of “involuntary” is compromised.
- The Corruption Dilemma: There is a tension between viewing the semiotic field as a “corruption” of a signal and viewing it as the “evolution” of that signal. The dialogue suggests it is both: a loss of original evolutionary function but a gain in social utility.
4. Areas for Further Exploration
- Cultural Neuroplasticity: How do specific cultural rituals (e.g., extreme spice eating or ritual scarification) physically alter the density or sensitivity of TRPV1 receptors over generations?
- The Ethics of the “Perfect Mask”: If empathy is a hackable biological reflex, what are the implications for a society where “performative suffering” can be used to garner resources or social capital?
- The Placebo of Semiotics: Can the “semiotic field” (the meaning we give to pain) act as a physiological analgesic, effectively shutting down the nociceptive signal before it reaches the cortex?
5. Conclusions on the Original Question
The original question asked if the transition from a biological absolute to a semiotic field is a “corruption of honesty” or if “biological honesty” is a social construct.
The synthesis suggests a third path: “Biological honesty” is a foundational myth used to anchor social meaning. While the chemical binding of capsaicin is a physical fact, it is never “honest” in a vacuum; it is always interpreted. Therefore, the transition into a semiotic field is not a “corruption” but the inevitable human process of meaning-making.
Ultimately, “honesty” in the human context is a performance that has been so thoroughly internalized that it bypasses the subject’s conscious awareness. The “biological absolute” is merely the stage upon which the social drama of suffering is performed; the two are not separate entities, but a single, integrated feedback loop of biology and culture.
Completed: 2026-02-20 20:12:53
| Total Time: 136.245s | Exchanges: 5 | Avg Exchange Time: 24.7088s |
Multi-Perspective Analysis Transcript
Subject: The Capsaicin Paradox: Biological Honesty vs. Social Signaling in Spicy Food Consumption
Perspectives: Biological/Evolutionary (The Plant and the Nervous System), Sociological/Cultural (Status, Identity, and Stereotypes), Commercial/Media (The Hot-Wing Industrial Complex), Psychological (The Individual Experience and Performance), Culinary/Authenticity (Gatekeeping and Flavor Complexity)
Consensus Threshold: 0.7
Context
Context Files and Prior Code
Context Files
/home/andrew/code/Science/scratch/2026-02-20-Sensory-Sociology/content.md
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
# The Capsaicin Paradox: Biological Honesty and the Social Poison of the Spicy Food Semiotic Field
The sensation of heat from a chili pepper is a biological absolute—a chemical interaction between capsaicin and the TRPV1 receptors that signals a warning of thermal pain. This is what we might call "biological honesty": the body reacting to a perceived threat with a predictable, physiological response. However, once this sensation enters the human social sphere, it is immediately transmuted into something far more complex. It becomes a "social poison," where the raw experience of pain is filtered through layers of cultural meaning, gendered performance, and status signaling. To understand this transformation, we must map the **Spicy Food Semiotic Field**—a conceptual space where the physiological reality of the burn collides with the symbolic weight of its consumption.
## The Biological Honesty of Heat
At its core, the capsaicinoid response is an evolutionary defense mechanism. Plants of the genus *Capsicum* developed capsaicin as a chemical deterrent against mammalian seed predators, whose molars would otherwise destroy the plant's reproductive potential. Birds, lacking the specific vanilloid receptors (TRPV1) sensitive to capsaicin, consume the fruit and disperse the seeds intact. This is the first layer of biological honesty: a clear, chemical "no" directed at the mammalian digestive system.
When a human bypasses this warning, the resulting sensation acts as a profound **embodiment anchor**. Unlike the subtle nuances of flavor, the heat of a habanero or a ghost pepper demands immediate, total interoceptive awareness. It forces the consciousness back into the physical frame, demanding attention to the mouth, the throat, and the rising internal temperature. In an era of digital abstraction, this "honest" pain provides a grounding reality that cannot be ignored or intellectualized away.
Physiologically, this heat serves several utilitarian functions. It acts as a built-in appetite throttle, increasing satiety and slowing the pace of consumption—a natural defense against overindulgence. Furthermore, its role as a vasodilator and irritant triggers a sinus-clearing mechanism, facilitating respiratory relief.
The transition from pain to pleasure—the "acquired taste"—is driven by the body's own compensatory mechanisms. As the TRPV1 receptors signal intense heat, the brain responds by releasing a flood of endorphins and dopamine to mitigate the perceived trauma. This creates the "endorphin loop," where the initial distress is followed by a mild euphoria. Over time, the brain learns to associate the burn not with damage, but with the subsequent neurochemical reward, transforming a biological warning into a sought-after culinary experience.
## The Social Layer and the Semiotic Field
The biological honesty of the capsaicin response—the involuntary flush, the beads of sweat, the dilated pupils—is precisely what makes it such fertile ground for social semiotics. Because these reactions are difficult to fake, they are treated as "honest signals" of an internal state. However, the moment these signals are observed by another, they are pulled into the **Semiotic Field**, where they are no longer just physiological events but symbols to be interpreted.
This is where the "Social Poison" of interpretive overfitting begins. In a social context, we rarely allow a physiological reaction to simply *be*; we over-interpret it, assigning intent and character to involuntary biological processes. The raw data of a sweating brow is overwritten by cultural scripts, leading to two primary, often conflicting, narratives.
### The Performance Hypothesis
The first narrative is the **Performance Hypothesis**, which views the consumption of extreme heat as a calculated display of status or belonging. In this framework, the visible struggle with spice is seen as a performative act—a "masochistic" display designed to signal toughness, worldliness, or membership in a specific subculture (such as the "chili-head" community). Here, the "honesty" of the biological response is ironically subverted; the very fact that the reaction is involuntary makes it the perfect stage for a performance of endurance. The consumer isn't just eating; they are demonstrating their ability to colonize their own biological pain for social capital.
### The Toughness Hypothesis
The second narrative is the **Toughness Hypothesis**, which interprets spice tolerance as a proxy for moral or physical character. In many cultures, the ability to "handle" heat is conflated with grit, resilience, and even masculinity. Under this lens, the person who does *not* sweat or flush is seen as possessing a superior constitution. The biological variation in TRPV1 receptor density—a matter of genetic luck—is transformed into a moral trait. To have a high tolerance is to be "strong"; to struggle is to be "weak." This hypothesis ignores the biological reality of desensitization and genetics, replacing it with a semiotic system where the tongue becomes a site of character judgment.
## The Sensory Semiotic Triad: Spicy, Sour, and Bitter
To fully appreciate the unique position of heat in the social hierarchy, we must compare it to other intense sensory experiences. We can conceptualize a **Sensory Semiotic Triad** consisting of the spicy, the sour, and the bitter. While all three involve a degree of "acquired" appreciation, they occupy vastly different roles in the theater of social interaction.
Spicy and sour sensations are inherently **public dramas**. Both trigger immediate, involuntary, and highly visible physiological responses—the "sour face" (puckering of the lips, squinting of the eyes) and the "spicy flush" (sweating, gasping, redness). These signals are ambiguous; they can represent either distress or intense pleasure, creating a semiotic gap that the observer must fill. This ambiguity is what allows them to be co-opted for performance. The "sour challenge" or the "hot wing challenge" relies on this visibility to turn a private sensation into a shared spectacle.
In contrast, bitterness remains largely a **private signal**. While a profound bitterness might cause a brief grimace, it lacks the sustained, escalating physical drama of heat or the sharp, contorting reflex of sourness. Bitterness is often a matter of internal preference—a "quiet" taste that one either enjoys or avoids. There is little social capital to be gained from "enduring" a bitter radicchio or a dark coffee in the same way one "conquers" a ghost pepper. Because the bodily response to bitterness is less legible to the observer and less prone to dramatic escalation, it rarely becomes a site of performative status. It is a preference, not a feat; a private dialogue between the palate and the brain, rather than a public broadcast of the body's resilience.
## Cultural Metastasis and Status Games
As the individual performance of spice consumption scales, it metastasizes into broader cultural tropes and identity politics. The semiotic field expands from the personal to the collective, where heat becomes a form of **cultural capital**. In this arena, the ability to navigate extreme spice is no longer just about personal toughness; it is a signal of worldliness and "authentic" engagement with the Other.
### Spice as Cultural Capital and Authenticity Policing
In the globalized culinary market, spice levels are often used as a metric for authenticity. This leads to a form of **authenticity policing**, where the "true" version of a dish is defined by its proximity to a perceived threshold of pain. Diners may demand "Thai spicy" or "Indian spicy" as a way to distance themselves from the "diluted" or "Westernized" versions of a cuisine. Here, the heat is not just a flavor profile but a gatekeeping mechanism. To consume the heat is to prove one's worthiness to participate in the culture, while the inability to do so is dismissed as a lack of culinary sophistication or a sheltered upbringing.
### The Sophistication Paradox
This leads to the **sophistication paradox**: the claim that a high tolerance for heat signals a more refined and adventurous palate. Proponents of this view argue that spice enhances the complexity of a dish, yet at extreme levels, capsaicin objectively obliterates flavor nuance by overwhelming the taste buds and triggering a pain response that drowns out subtle aromatics. The paradox lies in the fact that the "refined" palate is often celebrating the very thing that prevents it from tasting the food's finer details. The heat becomes a proxy for sophistication, even as it acts as a sensory blackout curtain.
### The Mayonnaise Proxy
Conversely, the absence of spice is weaponized through the **"mayonnaise" stereotype**. In contemporary social discourse, "mayonnaise" and "blandness" have become linguistic proxies for a specific type of identity—often associated with whiteness, lack of culture, or a fragile, unadventurous constitution. This stereotype functions as the inverse of the toughness hypothesis; if heat is grit, then mildness is weakness. By reducing complex cultural identities to their perceived tolerance for capsaicin, the semiotic field transforms a condiment into a tool for social categorization and identity-based derision. The "mayonnaise" label suggests a sensory sterility that is equated with a lack of historical or cultural depth, further entrenching the spicy food semiotic field as a primary site of modern social signaling.
## The Industrialization of Suffering
The final stage in the evolution of the spicy food semiotic field is its commercialization and media-driven escalation—what might be termed the **Hot-Wing Industrial Complex**. In this phase, the private biological struggle is commodified and transformed into a public spectacle. Media platforms, most notably YouTube series like *Hot Ones*, have pioneered a new genre of interview where the physiological distress of the subject is the primary draw. The "biological honesty" of the capsaicin response is leveraged as a tool for "authentic" celebrity exposure; the theory being that a person cannot maintain a PR-managed facade while their TRPV1 receptors are screaming.
This industrialization relies heavily on **Scoville-based marketing**, where the heat level of a product is no longer a culinary choice but a competitive metric. Sauces are marketed with names that evoke violence, death, or insanity, turning the grocery aisle into a gauntlet of performative bravery. This turns the act of eating into a "challenge"—a discrete event designed for social media documentation. The biological struggle is no longer a byproduct of a meal; it is the product itself.
Furthermore, this spectacle creates a new form of social cohesion: the **ritual of suffering together**. Whether in a viral "One Chip Challenge" or a group outing to a wing joint with a "wall of flame," the shared experience of intense, non-lethal pain acts as a powerful bonding mechanism. This is a secular, sensory ritual where the participants prove their mettle and their shared identity through a collective descent into physiological chaos. By turning the burn into a stage for performing identity, the Hot-Wing Industrial Complex ensures that the capsaicin paradox remains a central, if increasingly manufactured, pillar of modern social interaction.
## Conclusion: The Body as a Semiotic Surface
The journey from the TRPV1 receptor to the "Hot-Wing Industrial Complex" illustrates that the human body is never a closed biological system. It is a semiotic surface—a canvas upon which society projects its values of toughness, authenticity, and status. The "biological honesty" of a flush or a tear is immediately captured and recoded by the "social poison" of interpretation.
This machinery is not unique to capsaicin. We see identical patterns in the rise of extreme wellness rituals, such as the "cold plunge" or high-intensity interval training. In these contexts, the physiological shock—the "cold shock response"—is marketed and performed as a sign of superior discipline and mental fortitude. The shivering body, much like the sweating tongue, becomes a site of moral signaling. The raw data of the nervous system is consistently overwritten by the software of social expectation.
Ultimately, the capsaicin paradox reveals a fundamental human drive: the need to metabolize raw, chaotic sensation into structured, social meaning. We do not merely inhabit our bodies; we narrate them. By transforming a plant's defense mechanism into a cultural metric, we demonstrate that even our most visceral pains are never truly our own—they are always already part of the stories we tell about who we are and what we value.
Prior Code
Biological/Evolutionary (The Plant and the Nervous System) Perspective
This analysis examines the “Capsaicin Paradox” through the lens of the Biological/Evolutionary perspective, focusing specifically on the co-evolutionary struggle between the genus Capsicum and the mammalian nervous system.
1. The Biological/Evolutionary Analysis
A. The Plant’s Strategy: Directed Deterrence
From the perspective of the plant, capsaicin is a sophisticated secondary metabolite designed for Directed Deterrence. Evolutionarily, Capsicum plants faced a selective pressure: how to ensure seed dispersal while avoiding seed predation.
- The Mammalian Threat: Mammals possess molars that grind seeds, rendering them non-viable. Furthermore, mammalian digestive tracts are often too slow or chemically harsh for these specific seeds.
- The Avian Ally: Birds lack the TRPV1 (Transient Receptor Potential Vanilloid 1) receptor sensitivity to capsaicin. They swallow the fruits whole, and their rapid digestive transit leaves seeds intact and “packaged” in fertilizer.
- The Chemical “No”: Capsaicin is a molecular key that fits perfectly into the mammalian TRPV1 receptor—a receptor normally reserved for detecting damaging thermal heat (>43°C). By triggering this receptor, the plant “lies” to the mammalian nervous system, signaling a life-threatening burn where there is no actual tissue damage.
B. The Nervous System: The TRPV1 Interface and “Biological Honesty”
The human nervous system interprets capsaicin through the peripheral sensory neurons.
- The Mechanism of the Burn: Capsaicin lowers the activation threshold of TRPV1 receptors. At room temperature, the receptor “fires” as if the mouth is on fire. This triggers the autonomic nervous system: vasodilation (flushing), diaphoresis (sweating), and lacrimation (tears).
- The Endorphin Loop (Benign Masochism): The brain, receiving signals of intense pain, initiates a compensatory survival response. It floods the system with endorphins (endogenous opioids) and dopamine to mask the perceived trauma. This creates a neurochemical “high.”
- Neuroplasticity and Desensitization: With repeated exposure, the nervous system exhibits “functional desensitization.” The TRPV1-expressing neurons become less responsive (a process called “chili bleaching”). Evolutionarily, this allows humans to bypass the plant’s defense, effectively winning the chemical arms race through neural adaptation.
2. Key Considerations, Risks, and Opportunities
Key Considerations:
- Genetic Variability: There is significant polymorphic variation in TRPV1 receptor density among humans. What is “socially signaled” as toughness may simply be a biological lack of receptors (low sensitivity).
- The “Second Burn”: The nervous system’s interaction with capsaicin does not end at the mouth. TRPV1 receptors line the entire gastrointestinal tract, leading to increased motility and potential distress during elimination—a final evolutionary “parting gift” from the plant.
Risks:
- Sensory Overload (The Blackout): At extreme Scoville levels, the pain signal becomes so dominant that it causes “sensory crowding,” where the brain can no longer process flavor (gustatory) or aroma (olfactory) data.
- Vagal Overreaction: In rare cases, extreme capsaicin consumption can trigger a vasovagal response, leading to fainting or cardiac stress, as the body attempts to compensate for perceived extreme heat.
Opportunities:
- Therapeutic Desensitization: The same mechanism that allows “chili-heads” to eat ghost peppers is used in medical patches (e.g., Qutenza) to treat chronic neuropathy by “exhausting” pain receptors.
- Metabolic Thermogenesis: Capsaicin increases metabolic rate and fat oxidation. From an evolutionary standpoint, this “heat” is not just a sensation but a physiological reality that can aid in energy regulation.
3. Specific Insights and Recommendations
- Insight: The “Honesty” is Autonomic. The text refers to “biological honesty.” From a neurological perspective, this is because the response is mediated by the sympathetic nervous system. You cannot consciously tell your pores not to sweat or your capillaries not to dilate. This makes spicy food a unique “lie detector” for the human animal.
- Insight: Human Hijacking of Defense. Humans are the only mammals to systematically seek out capsaicin. We have transformed a deterrent into a reward by exploiting the brain’s own pain-mitigation circuitry (the endorphin loop).
- Recommendation for Analysis: When evaluating “spice tolerance,” one must distinguish between biological threshold (genetics), neural desensitization (habituation), and psychological framing (social signaling). A “tough” eater may simply have “bleached” their receptors through chronic exposure, meaning they are actually feeling less than the person who is struggling.
- Recommendation for Consumption: To maximize the “Endorphin Loop” without the “Sensory Blackout,” one should aim for the “threshold of perspiration”—the point where the autonomic nervous system just begins to react, signaling the release of opioids without overwhelming the gustatory cortex.
4. Confidence Rating
Confidence: 0.95 The biological mechanisms of capsaicin (TRPV1 interaction, directed deterrence, and the endorphin response) are well-established in ethnobotany and neurobiology. The interpretation of these biological facts as “honesty” vs. “social signaling” is a robust framework for understanding human-plant co-evolution.
Sociological/Cultural (Status, Identity, and Stereotypes) Perspective
This analysis examines The Capsaicin Paradox through the lens of Sociology and Cultural Studies, focusing on how the physiological act of consuming heat is transformed into a tool for status signaling, identity construction, and the reinforcement (or subversion) of stereotypes.
1. Sociological & Cultural Analysis
A. The Construction of “Sensory Capital”
In sociological terms, spice tolerance functions as a form of embodied cultural capital. Pierre Bourdieu’s theories suggest that taste is never neutral; it is a way to map social hierarchies.
- The Sophistication Paradox: By claiming to enjoy heat that “obliterates flavor,” the consumer signals a high-status “acquired taste.” This distinguishes the “refined” palate from the “vulgar” palate that seeks simple, unchallenging comfort.
- Globalism as Status: Navigating extreme spice levels (e.g., demanding “Thai spicy”) serves as a proxy for “worldliness.” It signals that the individual has traveled or possesses the cultural agility to engage with “authentic” non-Western traditions, distancing themselves from a perceived “sheltered” middle-class existence.
B. Gender and the Performance of “Grit”
The “Toughness Hypothesis” identifies spice consumption as a gendered performance.
- Masculinity and Stoicism: In many Western contexts, the ability to endure pain without visible distress is a hallmark of traditional masculinity. The “Hot-Wing Industrial Complex” (e.g., Hot Ones) commodifies this, turning a meal into a “challenge” or a “gauntlet.”
- Colonizing Pain: The text notes that consumers “colonize their own biological pain for social capital.” This reflects a broader sociological trend where the body is treated as a project to be mastered, disciplined, and displayed.
C. Identity Politics and the “Mayonnaise Proxy”
The use of spice—or the lack thereof—has become a potent linguistic tool in identity politics.
- Stereotyping the “Bland”: The “mayonnaise” stereotype functions as a sociological shorthand for whiteness, lack of culture, and fragility. It weaponizes sensory preference to categorize entire groups as “unadventurous” or “sterile.”
- Authenticity Policing: Spice levels are used to gatekeep cultural belonging. By defining “authentic” food through its pain threshold, the “chili-head” subculture creates a barrier to entry that prioritizes physiological endurance over historical or culinary understanding.
2. Key Considerations, Risks, and Opportunities
Key Considerations:
- The Biological-Moral Fallacy: A major sociological consideration is the conflation of a biological trait (TRPV1 receptor density) with a moral virtue (resilience). This creates a “hidden curriculum” in social dining where those with genetic sensitivity to capsaicin are coded as “weak.”
- The Ritual of Shared Suffering: Sociologically, the “One Chip Challenge” or group spicy-food outings function as secular rituals. They create “collective effervescence” (Durkheim), where the shared trauma of the burn bonds the group together against an external “threat.”
Risks:
- Reductionism: Reducing complex ethnic cuisines to their “heat” risks flattening diverse cultures into a single, painful caricature. This is a form of culinary orientalism, where the “Other” is valued only for the intensity of the sensation they provide.
- Performative Authenticity: The risk that the signal of eating spicy food becomes more important than the sustenance or the culture itself. This leads to “sensory blackout,” where the actual nuances of a culture’s cuisine are lost to the performative need for heat.
Opportunities:
- Subculture Formation: The “chili-head” community offers a unique space for cross-class and cross-cultural bonding based on a shared physical pursuit, potentially bypassing traditional social barriers.
- Deconstructing the “PR Facade”: As seen in media like Hot Ones, the “biological honesty” of spice can be used to humanize public figures, stripping away managed identities to reveal a raw, universal human reaction.
3. Specific Insights & Recommendations
- Insight: The “Mayonnaise” Reversal. Interestingly, the “mayonnaise” trope represents a rare instance where a “bland” dominant culture is mocked by using the sensory standards of marginalized or “global” cultures. It is a reversal of the traditional colonial gaze, where the “exotic” was once judged by Western standards of “refinement.”
- Insight: Spice as “Digital Grounding.” In an increasingly alienated, digital society, the “embodiment anchor” of capsaicin provides a rare moment of undeniable physical reality. The popularity of extreme spice may be a reaction to the “weightlessness” of modern life.
- Recommendation for Cultural Analysts: When observing food trends, distinguish between culinary appreciation (valuing the flavor profile of a chili) and semiotic performance (valuing the Scoville rating). The former is an engagement with culture; the latter is an engagement with status.
- Recommendation for Marketers: Move beyond “violence-based” marketing (e.g., “Mega-Death Sauce”). There is an opening for a “Post-Challenge” spice culture that focuses on the Endorphin Loop and the “afterglow” of spice, rather than just the initial “trauma.”
4. Confidence Rating
Confidence: 0.92 The analysis strongly aligns with established sociological theories (Bourdieu, Durkheim) and accurately deconstructs the semiotic cues provided in the source text. The connection between biological response and social signaling is a well-documented phenomenon in sensory sociology.
Commercial/Media (The Hot-Wing Industrial Complex) Perspective
Analysis: The Hot-Wing Industrial Complex (HWIC) Perspective
From the perspective of the Hot-Wing Industrial Complex (HWIC), the “Capsaicin Paradox” is not a philosophical dilemma but a highly efficient monetization engine. The transition from “biological honesty” to “social signaling” represents the exact point where a physiological reflex becomes a marketable commodity. In this view, the TRPV1 receptor is a consumer interface, and the resulting “social poison” is the primary driver of brand engagement and content virality.
1. Key Considerations: The Mechanics of the Market
- Authenticity as a Product: The HWIC views “biological honesty” as a tool for de-platforming the curated celebrity image. In media formats like Hot Ones, the capsaicin response is a “truth serum.” For the commercial side, this means the product’s value isn’t in its flavor profile, but in its ability to strip away a subject’s PR facade. The “sweat” is the ROI.
- The Scoville Arms Race: From a commercial standpoint, Scoville Heat Units (SHU) are not scientific measurements; they are performance metrics. High SHU ratings function like “horsepower” in the automotive industry—they provide a quantifiable basis for competition, comparison, and “toughness” signaling.
- The Gamification of Consumption: The “Performance Hypothesis” mentioned in the text is the foundation of the “Challenge” economy. By framing consumption as a “feat” or “conquest,” the HWIC transforms a one-time food purchase into a repeatable social media event (e.g., the “One Chip Challenge,” “Wall of Flame” rewards).
- The Sophistication Paradox as Market Segmentation: The HWIC leverages the “Sophistication Paradox” to create a premium tier of products. By marketing “artisanal” or “small-batch” extreme sauces, brands can capture the “connoisseur” demographic who wish to distance themselves from the “masochistic” wing-eater while still participating in the same high-heat signaling.
2. Risks: The “Burnout” Factor
- The Liability of “Biological Honesty”: As the industry pushes toward higher concentrations of capsaicinoids (extracts vs. whole peppers), the risk of genuine medical emergencies increases. The death of the “biological honesty” narrative occurs when the “non-lethal pain” becomes actual physical trauma, leading to litigation and platform bans (as seen with the withdrawal of the Paqui One Chip Challenge).
- Desensitization and Diminishing Returns: There is a physiological ceiling to the “Endorphin Loop.” As consumers build tolerance, the “spectacle” requires ever-increasing heat levels to produce the same visible reaction. This creates a “Red Queen” race where brands must innovate toward more extreme (and potentially less palatable) products just to maintain the same level of social signaling.
- The “Mayonnaise” Backlash: Over-reliance on the “toughness” narrative risks alienating the broader market. If spicy food is marketed solely as a test of grit, it becomes a niche “bro-culture” product, losing its appeal as a culinary staple.
3. Opportunities: Scaling the Sensation
- The “Ritual of Suffering” Kits: There is a massive opportunity in productizing the “shared descent into physiological chaos.” Selling “Home Challenge Kits” that include the sauce, the recording instructions, and the “merit badge” (stickers, t-shirts) allows the HWIC to decentralize the media spectacle.
- Cross-Vertical Integration: The “Sensory Semiotic Triad” (Spicy, Sour, Bitter) suggests room for expansion. The HWIC can expand into the “Extreme Sour” or “Extreme Bitter” markets using the same “Challenge” framework, though spicy remains the most “legible” for video-based media.
- The “Authenticity Policing” Premium: Brands can capitalize on the “Thai Spicy” vs. “Westernized” divide by offering “Origin-Specific” heat levels, charging a premium for “unfiltered” or “historically accurate” pain.
4. Strategic Recommendations
- Pivot from “Heat” to “Experience”: To avoid the Scoville ceiling, brands should market the narrative of the burn (e.g., “The 15-Minute Journey”) rather than just the peak SHU. This extends the “public drama” and provides more “content” for the consumer to share.
- Leverage the “Embodiment Anchor”: In an increasingly digital world, market extreme spice as a “Digital Detox” or a “Grounding Ritual.” Position the product as a way to “feel something real” in a world of abstraction.
- Institutionalize the “Wall of Flame”: Create digital “Walls of Fame” where consumers can upload their reaction videos directly to the brand’s ecosystem, turning the “Performance Hypothesis” into a self-sustaining marketing loop.
- Safety-First Branding: To mitigate liability, adopt “Extreme Sports” style waivers and clear “Safety Tiers.” This ironically increases the “toughness” signaling by making the product seem more dangerous than it is.
5. Confidence Rating
0.95 The analysis aligns perfectly with current market trends in the “eat-ertainment” sector and the documented success of media properties like First We Feast. The transition of food from “sustenance” to “spectacle” is a cornerstone of modern media-driven commerce.
Psychological (The Individual Experience and Performance) Perspective
This analysis examines The Capsaicin Paradox through the lens of Psychology, focusing on the individual’s internal experience, the construction of self-identity, and the mechanics of performance.
1. Psychological Analysis: The Individual Experience and Performance
From a psychological perspective, the consumption of capsaicin is a masterclass in the tension between interoception (the sense of the internal state of the body) and impression management (how we control others’ perceptions of us).
A. The “Embodiment Anchor” and Interoceptive Grounding
The text identifies capsaicin as a “biological honesty” mechanism. Psychologically, this functions as an intense grounding technique. In a state of “digital abstraction” or dissociation, the burning sensation forces the individual into a state of radical presence.
- The Flow State of Pain: Extreme heat demands total cognitive load. For the duration of the burn, the individual cannot ruminate on the past or worry about the future; they are locked in a “sensory now.” This can be psychologically cathartic, providing a temporary escape from complex emotional states through a simplified, singular physical focus.
B. Benign Masochism and the “Safe Threat”
The transition from pain to pleasure is best explained by Paul Rozin’s theory of Benign Masochism.
- Cognitive Appraisal: The individual’s brain receives “danger” signals from TRPV1 receptors, but the higher-order cognitive functions recognize there is no actual tissue damage.
- The Mastery Loop: The pleasure derived is not just neurochemical (endorphins); it is the psychological satisfaction of “tricking” the body. The individual experiences a sense of mastery over their own evolutionary hardware, enjoying the “rush” of a threat without the “risk” of the harm.
C. Identity Construction and the “Toughness” Schema
The “Toughness Hypothesis” mentioned in the text relates to Self-Schema Theory. If an individual views themselves as resilient, adventurous, or “gritty,” spicy food becomes a tool for self-verification.
- Internal Validation: “I am eating this ghost pepper; therefore, I am the type of person who can endure hardship.”
- The Moralization of Physiology: Individuals often commit the Fundamental Attribution Error regarding spice tolerance—attributing a biological/genetic trait (receptor density) to a character strength (willpower). This leads to an inflated sense of “moral” superiority in the high-tolerance individual.
D. Performance and the “Honest Signal”
Because the physiological responses (sweating, flushing) are autonomic and difficult to fake, they serve as “Honest Signals” in social psychology.
- The Vulnerability Paradox: By intentionally triggering a visible, “honest” distress response in a social setting (like Hot Ones), the individual performs a paradoxical act of “controlled vulnerability.” This builds trust and rapport with an audience because the subject appears “unmasked” by the pain, making their performance feel more authentic than a standard social interaction.
2. Key Considerations, Risks, and Opportunities
Key Considerations:
- Individual Differences in Interoception: Not everyone experiences the “embodiment anchor” the same way. For those with high anxiety or sensory processing sensitivities, the “biological honesty” of heat may feel like a traumatic intrusion rather than a grounding experience.
- The Desensitization Treadmill: Psychologically, the “acquired taste” requires escalating doses to achieve the same endorphin rush, leading to a “hedonic treadmill” where the individual must seek increasingly dangerous levels of heat to feel the same sense of mastery.
Risks:
- Performative Exhaustion: The “Social Poison” aspect suggests a risk where individuals override their body’s genuine “no” to satisfy a social script (masculinity, worldliness). This can lead to a dissociation from one’s own physical boundaries.
- Social Exclusion and “Blandness” Stigma: The weaponization of spice (the “Mayonnaise Proxy”) creates a psychological environment where dietary preference is conflated with cultural or intellectual deficiency, leading to “authenticity anxiety.”
Opportunities:
- Therapeutic Grounding: Controlled capsaicin exposure could be explored as a somatic tool for individuals struggling with dissociation or emotional numbing, providing a safe, time-limited way to “re-enter” the body.
- Prosocial Bonding: The “ritual of suffering together” creates Collective Effervescence. Sharing a painful but safe experience lowers social barriers and fosters rapid group cohesion.
3. Specific Insights and Recommendations
- Insight: The “Sophistication Paradox” is a Cognitive Bias. The belief that high heat equals a “refined palate” is often a form of compensatory conviction. The individual ignores the sensory “blackout” caused by pain to maintain the self-image of a connoisseur.
- Recommendation for Individuals: Practice Mindful Consumption. Distinguish between the “Endorphin Loop” (internal pleasure) and the “Performance Hypothesis” (external signaling). Ask: “Am I eating this for the grounding experience, or because I want to be seen as someone who can handle it?”
- Recommendation for Social Dynamics: Recognize that “Spice Tolerance” is a poor proxy for character. De-coupling physiological reactions from moral judgments can reduce the “Social Poison” of the semiotic field, allowing for a more inclusive culinary experience that doesn’t require “mettle” to enjoy.
4. Confidence Rating
0.92 The analysis is grounded in established psychological principles (Benign Masochism, Interoception, Self-Schema Theory, and Impression Management) and aligns well with the provided text’s exploration of the “Capsaicin Paradox.”
Culinary/Authenticity (Gatekeeping and Flavor Complexity) Perspective
This analysis examines The Capsaicin Paradox through the lens of Culinary Authenticity, Gatekeeping, and Flavor Complexity. From this perspective, the primary tension lies between the use of heat as a functional ingredient that enhances a dish’s profile and its use as a performative tool that obscures culinary craft.
1. Key Considerations: The Tension Between Craft and Performance
- The “Sensory Blackout” vs. Layered Complexity: The text identifies a “Sophistication Paradox” where extreme heat is equated with a refined palate, despite capsaicin’s ability to physically overwhelm the taste buds. From a culinary standpoint, heat should act as a flavor carrier or a structural element (like acid or salt). When heat crosses the threshold into “performance,” it triggers a sensory blackout, rendering the chef’s work on aromatics, fermentation, and balance irrelevant.
- Authenticity as a Moving Target: The “Thai Spicy” or “Indian Spicy” demand is a form of culinary gatekeeping. It assumes that the “truest” version of a culture’s food is its most aggressive version. This ignores the internal diversity of those cuisines, where many dishes are intentionally mild to provide contrast. Authenticity is thus reduced to a singular, painful metric rather than a holistic understanding of a region’s pantry.
- The “Mayonnaise” Stigma and the Devaluation of Subtlety: The weaponization of “blandness” (the Mayonnaise Proxy) creates a culinary environment where subtlety is mistaken for a lack of skill or culture. This puts pressure on chefs to over-spice dishes to avoid being labeled “unauthentic” or “Westernized,” potentially sacrificing the delicate balance of ingredients like lemongrass, galangal, or cumin.
2. Risks: The Erosion of Culinary Nuance
- The Scoville Arms Race: The “Hot-Wing Industrial Complex” risks turning dining into an endurance sport rather than a gastronomic experience. The risk here is the commodification of pain over the appreciation of the Capsicum fruit’s diverse flavor profiles (e.g., the fruitiness of a habanero, the smokiness of a chipotle, or the earthiness of an ancho).
- Alienation through Elitism: Gatekeeping via spice levels creates a “hierarchy of the tongue.” This can alienate diners who have a biological sensitivity to capsaicin (low TRPV1 threshold) but who might otherwise appreciate the complex spice blends of global cuisines. It turns a communal table into a site of “character judgment.”
- Monoculturalism of “Heat”: By focusing solely on capsaicin-induced heat, the culinary world risks ignoring other forms of “pungency” or “bite,” such as the numbing mā of Sichuan peppercorns, the nasal pungency of wasabi/horseradish, or the sharp bite of raw alliums, which offer complexity without the same “social poison” of performative endurance.
3. Opportunities: Reclaiming the Palate
- Functional Heat Education: There is an opportunity to shift the narrative from “How much heat can you take?” to “What does this heat do for the dish?” Chefs can lead this by explaining how heat cuts through fat (as in spicy pork belly) or how it brightens acidity.
- The “Mild Authenticity” Movement: Culinary influencers and chefs can push back against gatekeeping by highlighting “authentic” dishes that are intentionally low-heat. This validates the complexity of “bland” or “mild” profiles and dismantles the “Mayonnaise Proxy.”
- Varietal Appreciation: Moving beyond Scoville units to focus on the terroir of peppers. Just as wine is discussed via grape variety, peppers can be marketed for their specific notes (citrus, berry, tobacco), encouraging diners to seek flavor rather than just the “endorphin loop.”
4. Specific Recommendations and Insights
- For Chefs/Restaurateurs:
- Avoid the “Authenticity Trap.” Instead of labeling dishes as “Thai Spicy,” use descriptive language like “Aromatic Heat” or “Building Heat.”
- Offer “heat-adjusting” components on the side (chili oils, sambals) to allow the diner to find their own balance between the “biological honesty” of their palate and the “social signaling” of the table.
- For Food Critics/Media:
- De-emphasize the “challenge” aspect of spicy food.
- Critique the balance of heat. If a dish is so hot that the primary ingredients are indistinguishable, it should be viewed as a culinary failure of balance, regardless of its “authenticity.”
- Insight on the “Social Poison”: The most “authentic” culinary experience is one where the diner can actually taste the ingredients. When the “Social Poison” of performance takes over, the food ceases to be food and becomes a prop. The goal of a sophisticated palate should be discernment, not endurance.
5. Confidence Rating
0.95 The analysis strongly aligns with the provided text’s sociological framework while applying established culinary principles regarding flavor balance and the cultural politics of the modern dining scene.
Synthesis
This synthesis integrates five distinct perspectives—Biological, Sociological, Commercial, Psychological, and Culinary—to provide a unified conclusion on The Capsaicin Paradox.
1. Common Themes and Agreements
Across all analyses, three foundational themes emerge that define the human relationship with capsaicin:
- The Hijacked Defense (Biological to Psychological): There is universal agreement that capsaicin is an evolutionary deterrent (Directed Deterrence) designed to cause pain. Humans have uniquely “hijacked” this signal, transforming a survival-based “no” into a neurochemical “yes” through the Endorphin Loop and Benign Masochism. The brain recognizes the “safe threat,” allowing the individual to enjoy a physiological “high” without tissue damage.
- The Duality of the Signal (Honesty vs. Performance): Every perspective acknowledges that the body’s reaction to heat (sweating, flushing, lacrimation) is autonomically honest. Because these signals cannot be faked, they serve as a “truth serum.” However, this biological honesty is almost always co-opted for social signaling, where the ability to endure the “unmasking” of the burn is used to project toughness, worldliness, or cultural authenticity.
- The Escalation Treadmill (Desensitization to Commercialization): The Biological (neural bleaching), Psychological (hedonic treadmill), and Commercial (Scoville arms race) perspectives all agree that repeated exposure leads to diminishing returns. This necessitates ever-increasing heat levels to achieve the same internal “grounding” or external “spectacle,” driving the market toward extreme, and potentially hazardous, concentrations.
2. Key Conflicts and Tensions
While the perspectives agree on the mechanisms of the paradox, they conflict on the value of the experience:
- Flavor vs. Feat (Culinary vs. Commercial/Sociological): The Culinary perspective views extreme heat as a “sensory blackout” that destroys the nuance of a dish. Conversely, the Commercial and Sociological perspectives argue that in the context of “eat-ertainment,” the pain is the product. In these realms, the loss of flavor is a secondary concern to the successful performance of “grit.”
- Biological Luck vs. Moral Virtue (Biological vs. Psychological/Sociological): A significant tension exists between the reality of TRPV1 receptor density (genetic luck) and the social interpretation of spice tolerance. Society treats the ability to eat spicy food as a “moral” or “character” strength (resilience), whereas biology suggests it may simply be a lack of sensory hardware or “bleached” receptors.
- Authenticity vs. Caricature (Culinary vs. Sociological): The Sociological perspective notes that high heat is used as a proxy for “authentic” global culture (the “Mayonnaise Proxy”). The Culinary perspective counters that this is a form of “culinary orientalism” that ignores the subtle, mild, and complex dimensions of those same cuisines in favor of a singular, painful metric.
3. Overall Consensus Level
Consensus Rating: 0.92 There is a very high level of consensus regarding the existence and mechanics of the Capsaicin Paradox. All perspectives agree that spicy food consumption is a multi-layered event where a private biological reflex is transformed into a public social performance. The only minor divergence lies in the normative judgment of whether this transformation is a “sophisticated” evolution of taste or a “masochistic” degradation of culinary art.
4. Unified Recommendations
To navigate the Capsaicin Paradox effectively—whether as a consumer, creator, or critic—the following balanced approach is recommended:
- For Consumers: Seek “Interoceptive Grounding” over “Social Signaling.” To maximize the psychological benefits of the “Endorphin Loop” without the “Sensory Blackout,” aim for the “threshold of perspiration.” This is the point where the autonomic nervous system reacts, providing the “embodiment anchor” and neurochemical reward, without overwhelming the ability to taste and appreciate the food.
- For Media and Marketers: Pivot from “Violence” to “Journey.” The “Hot-Wing Industrial Complex” should move away from “lethality” branding (which carries high liability and diminishing returns) and toward the “narrative of the burn.” Marketing the “15-minute journey” or the “afterglow” of a specific pepper variety offers a more sustainable and premium “experience” than mere Scoville escalation.
- For Culinary Practitioners: Reclaim “Functional Heat.” Chefs should resist the “Authenticity Trap” that equates heat with quality. Heat should be used as a structural element (like acid) to cut through fat or brighten aromatics. Providing “heat-adjusting” components on the side allows diners to find their own balance between their biological threshold and the chef’s flavor profile.
- For Society: De-couple Physiology from Character. We should recognize that “spice tolerance” is a poor proxy for courage or cultural sophistication. Dismantling the “Mayonnaise Proxy” allows for a more inclusive culinary culture that values subtlety and biological diversity as much as it values the “ritual of shared suffering.”
Final Conclusion
The Capsaicin Paradox is the ultimate intersection of nature and nurture. It is a unique human achievement to have transformed a plant’s chemical “weapon” into a tool for self-mastery, social bonding, and commercial spectacle. While the “social poison” of performative toughness will likely persist, the most profound value of the chili pepper remains its ability to provide a moment of “biological honesty” in an increasingly abstracted world.
Persuasive Essay Generation Transcript
Started: 2026-02-20 20:57:52
Thesis: While the physiological response to capsaicin is an involuntary ‘honest signal’ of the body, its co-optation into the ‘Spicy Food Semiotic Field’ has transformed a biological warning into a social poison—a performative theater of status, ‘toughness,’ and commodified suffering that ultimately alienates us from genuine sensory experience.
Cover Image
Prompt:

Configuration
Persuasive Essay Generation
Thesis: While the physiological response to capsaicin is an involuntary ‘honest signal’ of the body, its co-optation into the ‘Spicy Food Semiotic Field’ has transformed a biological warning into a social poison—a performative theater of status, ‘toughness,’ and commodified suffering that ultimately alienates us from genuine sensory experience.
Configuration
- Target Audience: intellectually curious general public and cultural critics
- Tone: analytical and compelling
- Target Word Count: 1200
- Number of Arguments: 3
- Include Counterarguments: ✓
- Use Rhetorical Devices: ✓
- Include Evidence: ✓
- Use Analogies: ✓
- Call to Action: reflective
Started: 2026-02-20 20:58:08
Progress
Phase 1: Research & Outline
Analyzing thesis and creating essay structure…
1
2
3
Research Context
# /home/andrew/code/Science/scratch/2026-02-20-Sensory-Sociology/content.md
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
# The Capsaicin Paradox: Biological Honesty and the Social Poison of the Spicy Food Semiotic Field
The biological honesty of the capsaicin response—the involuntary flush, the beads of sweat, the dilated pupils—is precisely what makes it such fertile ground for social semiotics. Because these reactions are difficult to fake, they are treated as "honest signals" of an internal state. Yet this very treatment reveals a deeper philosophical move: what we call "biological honesty" is itself a form of **strategic essentialism**—a collective decision to treat the body's signals as ground truth so that we have a fixed reference point in an otherwise endlessly performative social world. The body's reactions are not *inherently* more truthful than any other sign; we *designate* them as such because a semiotic system requires an anchor, a place where interpretation agrees to stop. This anchoring is not a discovery about nature but a cultural commitment—one that quietly underwrites every social meaning we subsequently extract from the burn.
However, the moment these signals are observed by another, they are pulled into the **Semiotic Field**, where they are no longer just physiological events but symbols to be interpreted.
The sensation of heat from a chili pepper is a biological absolute—a chemical interaction between capsaicin and the TRPV1 receptors that signals a warning of thermal pain. This is what we might call "biological honesty": the body reacting to a perceived threat with a predictable, physiological response. However, once this sensation enters the human social sphere, it is immediately transmuted into something far more complex. It becomes a "social poison," where the raw experience of pain is filtered through layers of cultural meaning, gendered performance, and status signaling. To understand this transformation, we must map the **Spicy Food Semiotic Field**—a conceptual space where the physiological reality of the burn collides with the symbolic weight of its consumption.
## The Biological Honesty of Heat
At its core, the capsaicinoid response is an evolutionary defense mechanism—and a remarkably precise one. Plants of the genus *Capsicum* developed capsaicin not as a broad-spectrum toxin but as a strategy of **Directed Deterrence**: a molecular key shaped to fit one specific lock. Capsaicin binds to the TRPV1 vanilloid receptor—a thermosensor calibrated to fire at temperatures above 43°C—and activates it at room temperature. In effect, the molecule *lies* to the mammalian nervous system, convincing it that the mouth is being scalded when no thermal event is occurring. Birds, lacking the specific TRPV1 configuration sensitive to capsaicin, consume the fruit and disperse the seeds intact through their digestive tracts. The evolutionary elegance is striking: the plant does not poison indiscriminately but engineers a targeted sensory hallucination, exploiting the mammalian pain architecture to redirect its reproductive future through avian vectors. This is the first layer of biological honesty—though, as we shall see, even calling it "honesty" requires qualification, since the signal is, at the molecular level, a sophisticated deception.
When a human bypasses this warning, the resulting sensation acts as a profound **embodiment anchor**. Unlike the subtle nuances of flavor, the heat of a habanero or a ghost pepper demands immediate, total interoceptive awareness. It forces the consciousness back into the physical frame, demanding attention to the mouth, the throat, and the rising internal temperature. In an era of digital abstraction, this "honest" pain provides a grounding reality that cannot be ignored or intellectualized away. Yet even here, a crucial philosophical nuance intervenes—what we might call the **nociceptive-interpretive gap**. The TRPV1 receptor performs *nociception*: a straightforward chemical-to-electrical transduction, converting the presence of capsaicin into an ion channel event and a nerve impulse. But *pain*—the burning, the panic, the urge to reach for milk—is not the signal itself; it is the brain's *interpretation* of that signal, constructed through layers of context, memory, and expectation. Even at the most "biological" level, the body is not simply reporting; it is already narrating. The supposedly raw datum of the burn is, from its very first synapse, an act of meaning-making. This gap between transduction and experience is what makes the entire semiotic apparatus possible: if the signal were truly unmediated, there would be nothing for culture to rewrite.
Physiologically, this heat serves several utilitarian functions. It acts as a built-in appetite throttle, increasing satiety and slowing the pace of consumption—a natural defense against overindulgence. Furthermore, its role as a vasodilator and irritant triggers a sinus-clearing mechanism, facilitating respiratory relief.
The transition from pain to pleasure—the "acquired taste"—is driven by the body's own compensatory mechanisms. As the TRPV1 receptors signal intense heat, the brain responds by releasing a flood of endorphins and dopamine to mitigate the perceived trauma. This creates the "endorphin loop," where the initial distress is followed by a mild euphoria. Over time, the brain learns to associate the burn not with damage, but with the subsequent neurochemical reward, transforming a biological warning into a sought-after culinary experience. But there is a deeper physiological process at work beyond mere associative learning: **chili bleaching**, the functional desensitization of TRPV1-expressing neurons through repeated capsaicin exposure. With chronic stimulation, these neurons undergo a depletion of neuropeptides such as Substance P and, in some cases, a reversible degeneration of their peripheral terminals. The receptor doesn't "toughen up"; it is effectively silenced. What society reads as "toughness"—the stoic consumption of a Carolina Reaper without visible distress—may simply be the result of bleached receptors, a neurological quiet mistaken for a moral quality. And the body's encounter with capsaicin does not end at the palate. TRPV1 receptors line the entire gastrointestinal tract, from esophagus to rectum, and capsaicin activates them at every station along the way. This **"second burn"**—the intestinal cramping, the unmistakable rectal sting hours later—is the plant's evolutionary parting gift, a final reminder that the directed deterrence strategy extends well beyond the mouth. It is a sensation rarely discussed in polite company, yet it completes the biological circuit: the mammalian body is warned at entry *and* at exit, bookending the digestive journey with the same molecular lie.
### Benign Masochism and the Psychology of the Acquired Taste
Yet the endorphin loop and receptor desensitization, powerful as they are, do not fully explain why humans *seek out* the burn in the first place. After all, the body produces compensatory endorphins in response to many forms of pain—stubbing a toe, touching a hot stove—without those experiences becoming objects of craving. The missing piece is not neurochemical but cognitive, and it was most precisely articulated by the psychologist Paul Rozin, who coined the term **"benign masochism"** to describe the peculiar human capacity to derive pleasure from sensations that the body's alarm systems register as threats. Rozin's insight is that the pleasure of capsaicin is not merely the endorphin afterglow that follows the pain; it is the *simultaneous awareness* that the pain is real and the danger is not. The higher-order cortex—the evaluative, contextualizing brain—monitors the TRPV1 alarm and performs a rapid override: *the mouth is screaming "fire," but there is no fire; the nociceptive system is reporting tissue damage, but no tissue is being damaged.* It is this cognitive gap between the body's conviction and the mind's knowledge that generates the characteristic thrill. The pleasure derives, in significant part, from the psychological satisfaction of having *tricked one's own evolutionary hardware*—a felt sense of mastery over the body's most ancient and authoritative warning system. The eater is not merely enduring pain; they are staging a private rebellion against the tyranny of their own reflexes, and *winning*.
What Rozin identified, in effect, is the structure of a **"safe threat"**—an experience that provides all the physiological markers of a genuine biological crisis (the racing heart, the sweating, the flood of stress hormones) without any of the biological cost. This is the same architecture that underlies the appeal of roller coasters, horror films, and skydiving: the body is fully mobilized for an emergency that the mind knows will not arrive. But capsaicin occupies a unique position within this category, because the threat is not external and environmental but *internal and somatic*—the danger signal originates from inside the body itself, from the very tissue of the mouth, making the override feel more intimate, more like a genuine contest between self and self. This psychological dimension is the critical bridge between the biological story told above and the social story that follows. The endorphin loop explains how pain becomes tolerable; benign masochism explains how it becomes *desirable*. And it is precisely at the moment when the individual discovers this private pleasure—this quiet triumph of cognition over nociception, of the knowing mind over the screaming body—that the experience becomes available for social interpretation. The internal mastery must be *performed* to be *witnessed*, and once it is witnessed, it enters the semiotic field. The private rebellion becomes a public signal. The body's honest alarm, already overridden by the individual's own cognition, is now ready to be overwritten again—this time by the collective scripts of culture, status, and identity.
## The Social Layer and the Semiotic Field
This is where the "Social Poison" of interpretive overfitting begins. In a social context, we rarely allow a physiological reaction to simply *be*; we over-interpret it, assigning intent and character to involuntary biological processes. The raw data of a sweating brow is overwritten by cultural scripts, leading to two primary, often conflicting, narratives.
### The Performance Hypothesis
The first narrative is the **Performance Hypothesis**, which views the consumption of extreme heat as a calculated display of status or belonging. In this framework, the visible struggle with spice is seen as a performative act—a "masochistic" display designed to signal toughness, worldliness, or membership in a specific subculture (such as the "chili-head" community). Here, the "honesty" of the biological response is ironically subverted; the very fact that the reaction is involuntary makes it the perfect stage for a performance of endurance. The consumer isn't just eating; they are demonstrating their ability to colonize their own biological pain for social capital.
What gives this performance its peculiar credibility is a dynamic that biologists call the **Handicap Principle** and game theorists call **costly signaling**: a signal is trustworthy precisely *because* it is expensive to produce. A peacock's tail is convincing evidence of genetic fitness because growing and carrying it is a genuine metabolic burden; a bluff would collapse under the weight. The spicy-food performance operates on the same logic. The visible struggle—the flushed skin, the watering eyes, the involuntary gasp—is not a flaw in the display; it *is* the display. It works as a status signal precisely because it hurts. The pain is the price of admission, and only those who value the resulting social capital enough to pay it, or who possess a naturally high capsaicin tolerance that lowers the cost, will sit down at the table and order the Carolina Reaper. Someone who merely *claims* to love extreme heat but blanches at the first bite gains nothing; the body's involuntary betrayal exposes the bluff instantly. This is the deep irony at the heart of the Performance Hypothesis: the biological honesty of the capsaicin response—the very thing that seems to undercut a polished social performance—is what makes the performance credible in the first place. Because the cost cannot be faked, the signal cannot be counterfeited. The sweating brow is not a failure of composure; it is the receipt that proves the toll was paid.
### The Toughness Hypothesis
The second narrative is the **Toughness Hypothesis**, which interprets spice tolerance as a proxy for moral or physical character. In many cultures, the ability to "handle" heat is conflated with grit, resilience, and even masculinity. Under this lens, the person who does *not* sweat or flush is seen as possessing a superior constitution. The biological variation in TRPV1 receptor density—a matter of genetic luck—is transformed into a moral trait. To have a high tolerance is to be "strong"; to struggle is to be "weak." This hypothesis ignores the biological reality of desensitization and genetics, replacing it with a semiotic system where the tongue becomes a site of character judgment.
From a psychological standpoint, this conflation of tolerance with character is a textbook instance of the **Fundamental Attribution Error**—the well-documented tendency to attribute others' behavior to stable dispositional traits (courage, weakness, discipline) while underweighting the situational and biological variables that actually explain the behavior. When an observer watches someone eat a scorpion pepper without flinching and concludes that this person is *tough*, they are performing precisely the attribution error that Lee Ross identified in 1977: mistaking a situational outcome—one shaped by prior exposure history, cultural habituation, and, crucially, genetic endowment—for a window into the person's character. The error is compounded by the fact that the biological substrate in question is not uniform across the population. The *TRPV1* gene exhibits significant **polymorphic variation**: single-nucleotide polymorphisms (SNPs) in and around the TRPV1 locus alter receptor expression levels, ion-channel sensitivity, and downstream signal amplification. Some individuals are born with a sparse distribution of functional TRPV1 receptors in their oral mucosa; others carry variants that produce receptors with a higher activation threshold, requiring more capsaicin to trigger the same nociceptive cascade. What society codes as "strength"—the calm, unflushed consumption of extreme heat—may therefore be, in the most literal sense, a **partial absence of sensory hardware**: not a triumph of will over pain, but a quieter peripheral signal that never reached the volume the observer imagines was heroically suppressed. Conversely, the person who gasps and reaches for milk may possess a denser receptor field or a more sensitive channel variant—a richer sensory apparatus, not a weaker constitution. The Toughness Hypothesis thus performs a double inversion: it reads a genetic variable as a moral constant, and it rewards sensory *deficit* as though it were psychological *surplus*. By mapping the Fundamental Attribution Error onto the biology of TRPV1 polymorphism, we can see that the entire edifice of spice-as-character-test rests on a failure to distinguish between what the body *is* and what the person *chooses*—between the hand one is dealt and the way one plays it.
## The Sensory Semiotic Triad: Spicy, Sour, and Bitter
To fully appreciate the unique position of heat in the social hierarchy, we must compare it to other intense sensory experiences. We can conceptualize a **Sensory Semiotic Triad** consisting of the spicy, the sour, and the bitter. While all three involve a degree of "acquired" appreciation, they occupy vastly different roles in the theater of social interaction.
Spicy and sour sensations are inherently **public dramas**. Both trigger immediate, involuntary, and highly visible physiological responses—the "sour face" (puckering of the lips, squinting of the eyes) and the "spicy flush" (sweating, gasping, redness). These signals are ambiguous; they can represent either distress or intense pleasure, creating a semiotic gap that the observer must fill. Crucially, these signals are what semioticians in the Peircean tradition would call **polysemic**—they carry multiple, competing meanings simultaneously. A flushed face and watering eyes could signify genuine agony, masochistic pleasure, performative bravado, or the threshold moment between surrender and triumph. The observer cannot determine which meaning is "correct" from the signal alone, and it is precisely this **semiotic ambiguity** that serves as the engine of social co-optation. Because the sign underdetermines its meaning, it *invites* interpretation—and where interpretation is invited, social narratives rush in. The "sour challenge" or the "hot wing challenge" relies on this polysemic visibility to turn a private sensation into a shared spectacle, a text that the audience actively co-authors.
In contrast, bitterness remains largely a **private signal**. While a profound bitterness might cause a brief grimace, it lacks the sustained, escalating physical drama of heat or the sharp, contorting reflex of sourness—there is no progressive arc of visible suffering, no crescendo for an audience to track. More fundamentally, the bitter response is **monosemic**: it means one thing—*rejection*. The grimace at a bitter compound is read, almost universally, as aversion. There is no interpretive gap, no ambiguity about whether the person is suffering beautifully or performing resilience. The sign arrives pre-interpreted, and where there is no interpretive gap, there is no space for social narrative to colonize. A polysemic signal is an open text; a monosemic signal is a closed one. No one watches someone sip an intensely bitter espresso and wonders, *"Is she in pain or ecstasy? Is this a feat of endurance?"*—the grammar of the response simply does not pose the question.
Bitterness is therefore processed as a matter of internal preference—a "quiet" taste that one either enjoys or avoids, a **private dialogue between the palate and the brain** rather than a public broadcast of the body's resilience. There is little social capital to be gained from "enduring" a bitter radicchio or a dark coffee in the same way one "conquers" a ghost pepper, because the bodily response to bitterness is neither legible enough to be read at a distance nor ambiguous enough to sustain competing interpretations. It is a preference, not a feat. The Triad thus reveals a principle: **the social life of a sensation is proportional to the polysemy of its signal**. Spicy and sour sensations become theaters of identity precisely because their signs are open to multiple readings; bitterness remains a private affair precisely because its sign is closed. The engine of social co-optation is not intensity alone—it is interpretive *possibility*.
## Cultural Metastasis and Status Games
As the individual performance of spice consumption scales, it metastasizes into broader cultural tropes and identity politics. The semiotic field expands from the personal to the collective, where heat becomes a form of **cultural capital**. In this arena, the ability to navigate extreme spice is no longer just about personal toughness; it is a signal of worldliness and "authentic" engagement with the Other.
### Spice as Cultural Capital and Authenticity Policing
In the globalized culinary market, spice levels are often used as a metric for authenticity. This leads to a form of **authenticity policing**, where the "true" version of a dish is defined by its proximity to a perceived threshold of pain. Diners may demand "Thai spicy" or "Indian spicy" as a way to distance themselves from the "diluted" or "Westernized" versions of a cuisine. Here, the heat is not just a flavor profile but a gatekeeping mechanism. To consume the heat is to prove one's worthiness to participate in the culture, while the inability to do so is dismissed as a lack of culinary sophistication or a sheltered upbringing.
Yet this gatekeeping mechanism carries a deeper distortion that culinary scholars have identified as **culinary orientalism**: the reduction of an entire cuisine's vast internal diversity to a single painful metric. To demand "Thai spicy" is to flatten a tradition in which many canonical dishes—the coconut-milk richness of *tom kha gai*, the delicate balance of *pad see ew*, the intentional mildness of a *kanom jeen* with mild fish curry—are designed to be gentle precisely *because* they provide contrast and balance within a meal structured around complementary sensations. Thai cuisine, like most complex culinary traditions, is an architecture of counterpoints: hot against cool, rich against sharp, intense against mild. To define its "authenticity" by its maximum capsaicin output is to mistake one voice in a choir for the entire performance—and, in doing so, to erase the compositional intelligence that makes the tradition worth celebrating in the first place. The semiotic field does not merely distort social dynamics; it flattens the very cuisines it claims to honor.
The narrowing runs deeper still. The semiotic field's fixation on capsaicin-based heat as *the* metric of spice courage ignores the fact that pungency itself is a diverse sensory landscape. The numbing, electric *mā* (麻) of Sichuan peppercorns—produced by hydroxy-alpha-sanshool acting on mechanoreceptors rather than thermoreceptors—offers a form of intensity that is neurologically distinct from the capsaicin burn: it tingles, buzzes, and temporarily rewires the mouth's sense of touch rather than its sense of temperature. The sharp nasal pungency of wasabi and horseradish, mediated by allyl isothiocyanate activating TRPA1 receptors in the sinuses, produces a fleeting, almost cerebral flash of pain that dissipates in seconds—a sensation with no sustained arc of visible suffering and therefore little utility for the endurance performances the semiotic field rewards. These alternative pungencies offer extraordinary complexity and cultural depth, yet they remain marginal to the dominant spice discourse precisely because they do not produce the prolonged, legible, polysemic spectacle that capsaicin does. The semiotic field, in other words, has not merely co-opted heat as a social signal; it has selected for a *specific kind* of heat—the kind that best serves performative endurance—and in doing so, has narrowed the sensory landscape even as it claims to celebrate adventurousness.
### The Sophistication Paradox
This leads to the **sophistication paradox**: the claim that a high tolerance for heat signals a more refined and adventurous palate. Proponents of this view argue that spice enhances the complexity of a dish, yet at extreme levels, capsaicin objectively obliterates flavor nuance by overwhelming the taste buds and triggering a pain response that drowns out subtle aromatics. The paradox lies in the fact that the "refined" palate is often celebrating the very thing that prevents it from tasting the food's finer details. The heat becomes a proxy for sophistication, even as it acts as a **sensory blackout curtain**—and the claim that extreme spice *enhances* one's appreciation of a dish is, as a culinary analogy makes vivid, like saying you appreciate music more because you listen to it at 120 decibels. At that volume, the subtlety of the composition is not revealed but annihilated; what remains is not discernment but endurance. Game theory sharpens the absurdity further. Within the spicy food semiotic field, social capital is often valued more than the primary utility of the food itself, and the sophistication paradox is the point where this priority structure becomes fully visible. The consumer who orders the hottest item on the menu is executing a strategic trade-off: sacrificing the **utility of flavor**—the very thing that ostensibly defines culinary sophistication—in order to gain the **utility of status**. This is not a hidden cost; it is the entire mechanism. The diner pays in destroyed nuance and collects in social capital. Yet the sophistication claim requires that this trade-off remain unacknowledged, because the moment it is made explicit, the paradox collapses into plain contradiction: one cannot simultaneously claim to be maximizing gustatory appreciation while pursuing a strategy that systematically minimizes it. The "refined palate" is, in game-theoretic terms, a player who has optimized for a payoff (status) that is structurally incompatible with the payoff they advertise (taste)—and the sensory blackout curtain is not an unfortunate side effect of their strategy but its operative mechanism.
### The Mayonnaise Proxy
Conversely, the absence of spice is weaponized through the **"mayonnaise" stereotype**. In contemporary social discourse, "mayonnaise" and "blandness" have become linguistic proxies for a specific type of identity—often associated with whiteness, lack of culture, or a fragile, unadventurous constitution. This stereotype functions as the inverse of the toughness hypothesis; if heat is grit, then mildness is weakness. By reducing complex cultural identities to their perceived tolerance for capsaicin, the semiotic field transforms a condiment into a tool for social categorization and identity-based derision. The "mayonnaise" label suggests a sensory sterility that is equated with a lack of historical or cultural depth, further entrenching the spicy food semiotic field as a primary site of modern social signaling.
## The Industrialization of Suffering
The final stage in the evolution of the spicy food semiotic field is its commercialization and media-driven escalation—what might be termed the **Hot-Wing Industrial Complex**. In this phase, the private biological struggle is commodified and transformed into a public spectacle. Media platforms, most notably YouTube series like *Hot Ones*, have pioneered a new genre of interview where the physiological distress of the subject is the primary draw. The "biological honesty" of the capsaicin response is leveraged as a tool for "authentic" celebrity exposure; the theory being that a person cannot maintain a PR-managed facade while their TRPV1 receptors are screaming.
This industrialization relies heavily on **Scoville-based marketing**, where the heat level of a product is no longer a culinary choice but a competitive metric. Sauces are marketed with names that evoke violence, death, or insanity, turning the grocery aisle into a gauntlet of performative bravery. This turns the act of eating into a "challenge"—a discrete event designed for social media documentation. The biological struggle is no longer a byproduct of a meal; it is the product itself. Game theory reveals the structural logic driving this escalation. The Mayonnaise Proxy penalty described above—the social cost of being seen to prefer mild food—functions as a powerful **negative payoff** that reshapes the entire decision landscape. When the stigma attached to choosing mildness is sufficiently severe, the semiotic field collapses into what game theorists call a **Pooling Equilibrium**: a state in which all players, regardless of their private type (their actual capsaicin tolerance or preference), choose the same publicly visible action—extreme heat—because the cost of revealing oneself as mild-preferring exceeds the physical cost of enduring the burn. High-tolerance and low-tolerance individuals alike converge on the same fiery order, not because they share a preference but because the signaling environment has made the alternative socially untenable. The menu ceases to be a space of genuine choice and becomes a one-dimensional race to the top of the Scoville scale, where the only "safe" move is to match or exceed the group's current threshold.
This pooling dynamic generates a **Prisoner's Dilemma** at the heart of every shared meal. Consider two diners at a table: each might privately prefer a milder dish—one that would allow them to taste the food's actual flavor profile and avoid the gastrointestinal aftermath. Yet each knows that choosing mildness while the other chooses heat will trigger the Mayonnaise Proxy penalty, coding them as the "weak" one in the pair. The dominant strategy for both players, therefore, is to choose spicy—not because it maximizes their individual gustatory utility, but because it minimizes their social risk. The result is a **Pareto-inferior equilibrium**: both diners end up suffering more than necessary, consuming food that overwhelms their palates and punishes their digestive tracts, when a world in which both chose mildly would have left each of them better off on every dimension except the one that matters in the semiotic field—status. The tragedy is structural, not personal. No single diner can unilaterally defect to mildness without bearing the full weight of the social penalty, and so the equilibrium holds, meal after meal, even though every participant in it would prefer a different world.
The commercial arm of the Hot-Wing Industrial Complex is locked into a parallel escalation that evolutionary biologists would recognize as a **Red Queen race**—the dynamic in which organisms must constantly evolve merely to maintain their current fitness relative to co-evolving competitors. As the pooling equilibrium drives consumers toward ever-higher Scoville counts, and as repeated exposure triggers the chili bleaching and TRPV1 desensitization described earlier, the spectacle's impact erodes. A sauce that once produced dramatic, camera-ready suffering becomes, through habituation, merely uncomfortable—and a merely uncomfortable sauce generates no social media content, no viral moments, no signaling value. Brands must therefore innovate relentlessly toward more extreme formulations—higher capsaicinoid concentrations, novel cultivars like Pepper X and Apollo, synthetic capsaicin analogs—not to *increase* the level of social signaling their products provide, but simply to *maintain* it against the rising baseline of consumer tolerance. They are running to stand still. The result is an arms race between the human nervous system's capacity for adaptation and the food industry's capacity for chemical escalation, each driving the other forward in a feedback loop that has no natural resting point. The Scoville number on the label is not a measure of flavor; it is an index of how far the race has run.
Furthermore, this spectacle creates a new form of social cohesion: the **ritual of suffering together**. Whether in a viral "One Chip Challenge" or a group outing to a wing joint with a "wall of flame," the shared experience of intense, non-lethal pain acts as a powerful bonding mechanism. This is a secular, sensory ritual where the participants prove their mettle and their shared identity through a collective descent into physiological chaos. The pooling equilibrium ensures that the ritual's entry price keeps rising—what counted as a credible performance of toughness five years ago is now the table stakes, and the Prisoner's Dilemma ensures that no individual participant can opt for a gentler experience without defecting from the group's implicit social contract. By turning the burn into a stage for performing identity, and by locking both consumers and producers into interlocking escalation dynamics—the former trapped in a Pareto-inferior social equilibrium, the latter sprinting on the Red Queen's treadmill—the Hot-Wing Industrial Complex ensures that the capsaicin paradox remains a central, if increasingly manufactured and increasingly painful, pillar of modern social interaction.
## Conclusion: The Body as a Semiotic Surface
The journey from the TRPV1 receptor to the "Hot-Wing Industrial Complex" illustrates that the human body is never a closed biological system. It is a semiotic surface—a canvas upon which society projects its values of toughness, authenticity, and status. The "biological honesty" of a flush or a tear is immediately captured and recoded by the "social poison" of interpretation.
What this entire apparatus amounts to is a **domestication of the absolute**. The capsaicin response begins as a biological absolute—a raw, unmediated chemical scream from the TRPV1 receptor, a signal that evolved to mean one thing and one thing only: *danger, stop, do not eat this*. Yet we take this absolute and domesticate it, breaking it like a wild animal to the harness of social utility. We train it to mean *toughness*, *authenticity*, *worldliness*, *belonging*. We build elaborate semiotic architectures—the Performance Hypothesis, the Toughness Hypothesis, the Sophistication Paradox, the Hot-Wing Industrial Complex—all of which are, at bottom, cultural infrastructure for converting a biological emergency signal into a legible social currency. The "honesty" we attribute to the body's involuntary response is not a property we discover in nature; it is a cultural layer we apply, a domesticating frame that makes sense of our peculiar desire to suffer for pleasure, for status, for the feeling of being fully alive in a body we can narrate to others. We do not honor the biological signal by reading it as honest; we *tame* it by doing so, rendering it safe enough to circulate in the economy of social meaning.
But domestication has a boundary, and that boundary reveals something essential about the entire enterprise. There exists a threshold—what we might call the point of **semiotic collapse**—where pain becomes so overwhelming that the social field dissolves entirely. Elaine Scarry, in *The Body in Pain* (1985), argued that intense physical pain is fundamentally *world-destroying*: it annihilates language, unmakes the sufferer's capacity for symbolic thought, and reduces the human subject to pre-linguistic cries that carry no social meaning whatsoever. At the extreme end of the capsaicin spectrum—the point where a Carolina Reaper or a concentrated extract overwhelms not just the palate but the entire nervous system—we approach this threshold. The sufferer no longer performs toughness or signals authenticity; they simply *hurt*, in a way that is illegible to any semiotic framework. The flush is no longer a polysemic sign open to competing interpretations; it is the body in revolt, beyond narrative, beyond performance, beyond meaning. Language fails. The groan that escapes is not a social signal but a biological reversion—the organism stripped of its cultural clothing, returned to the pre-symbolic animal cry that no audience can decode as anything other than what it is.
The entire spicy food semiotic field, then, operates in the narrow space between two poles: the **embodiment anchor**—the point where capsaicin forces consciousness back into the body and makes the flesh available for social narration—and the **threshold of semiotic collapse**—the point where pain overwhelms all narration and the body ceases to be a legible text. Every hot wing challenge, every "Thai spicy" request, every Scoville-escalation arms race is a **controlled flirtation with the biological real**: an approach toward the abyss of unsymbolizable pain that never quite tips over the edge. The social utility of the burn depends entirely on maintaining this balance—enough pain to produce the honest, costly signal that the semiotic field requires, but not so much that the signal dissolves into the formless scream that no culture can metabolize. The domestication of the absolute is, in this light, an act of extraordinary precision: we must bring the wild animal close enough to feel its heat, but not so close that it devours us.
This machinery is not unique to capsaicin. We see identical patterns in the rise of extreme wellness rituals, such as the "cold plunge" or high-intensity interval training. In these contexts, the physiological shock—the "cold shock response"—is marketed and performed as a sign of superior discipline and mental fortitude. The shivering body, much like the sweating tongue, becomes a site of moral signaling. The raw data of the nervous system is consistently overwritten by the software of social expectation. And in each case, the same two boundaries apply: the experience must be intense enough to anchor the body as a legible semiotic surface, but not so intense that it crosses into the territory Scarry mapped—the zone where pain unmakes the world and the social field goes dark.
Ultimately, the capsaicin paradox reveals a fundamental human drive: the need to metabolize raw, chaotic sensation into structured, social meaning. We do not merely inhabit our bodies; we narrate them. But the narration is always precarious, always conducted at the edge of a silence that would swallow it whole. By transforming a plant's defense mechanism into a cultural metric, we demonstrate that even our most visceral pains are never truly our own—they are always already part of the stories we tell about who we are and what we value. Yet the stories hold only so long as the pain remains domesticated. Beyond the last Scoville unit that language can metabolize lies the biological real—mute, absolute, and indifferent to every meaning we have tried to make it carry.
1
## Essay Outline
The Spicy Food Semiotic Field: From Biological Honesty to Social Poison
Hook
Describe the visceral, high-definition spectacle of a “Hot Ones” interview or a “One Chip Challenge” video—the streaming sweat, the panicked gasping, and the dilated pupils—contrasting the physical agony of the participant with the rapturous entertainment of the audience.
Background
Capsaicin, the active component in chili peppers, evolved as a “directed deterrence” mechanism—a molecular lie that tricks mammalian TRPV1 receptors into sensing a thermal burn where none exists. Historically, humans integrated this “burn” into culinary traditions for preservation and flavor. However, in the digital age, the biological response has been decoupled from nutrition. We have entered the “Spicy Food Semiotic Field,” where the involuntary physiological reactions (the “honest signals” of the body) are no longer private sensations but public currency. In a world of curated personas, the “unfakeable” nature of a capsaicin breakdown has become a prized commodity.
Thesis Statement
While the physiological response to capsaicin is an involuntary “honest signal” of the body, its co-optation into the “Spicy Food Semiotic Field” has transformed a biological warning into a social poison—a performative theater of status, “toughness,” and commodified suffering that ultimately alienates us from genuine sensory experience.
Main Arguments
Argument 1: We treat the body’s involuntary reaction to heat as a “ground truth” to anchor social meaning in an increasingly performative world.
Supporting Points:
- The Involuntary Signal: Unlike a smile or a gesture, the “capsaicin flush” and lacrimation (tearing) cannot be easily faked, making them “honest signals” in biological terms.
- Strategic Essentialism: We collectively decide to treat these signals as the “real” person emerging from behind the mask of social etiquette.
- The Semiotic Anchor: By designating the body’s pain as “truth,” we create a fixed reference point that allows us to judge a person’s “character” or “grit.”
Evidence Types: Evolutionary biology (TRPV1 receptor function), Sociological theories of ‘Strategic Essentialism’, Analysis of ‘Hot Ones’ as a ‘truth-seeking’ interview format
Rhetorical Approach: Logos/Ethos
Est. Words: 250
Argument 2: The “Spicy Food Semiotic Field” weaponizes pain to reinforce hierarchies of gender, endurance, and “masculine” stoicism.
Supporting Points:
- Gendered Performance: The “chili-head” subculture often mirrors “frontier masculinity,” where consuming pain is equated with virility and dominance.
- The Theater of Stoicism: The goal is no longer to enjoy the food, but to maintain a “poker face” while the body screams, turning a meal into a trial by ordeal.
- Status Signaling: High-Scoville consumption acts as a “costly signal”—a way to demonstrate superior constitution or “hardiness” to a social group.
Evidence Types: Analysis of marketing language for ‘extreme’ hot sauces, Social media ‘challenges’, Historical parallels to rites of passage
Rhetorical Approach: Pathos/Logos
Est. Words: 250
Argument 3: The industrialization of extreme spice (the “Scoville Arms Race”) destroys the nuance of flavor, alienating the consumer from the actual culinary experience.
Supporting Points:
- The Scoville Arms Race: The pursuit of pure capsaicin extracts prioritizes “heat units” over gastronomic complexity, turning food into a delivery system for a drug-like hit.
- The Spectacle of the “Social Poison”: When pain becomes a product to be sold (e.g., the “One Chip Challenge”), the consumer is no longer a diner but a performer in a commodified circus.
- Sensory Erasure: Extreme heat overwhelms the palate, effectively “blinding” the taster to the actual ingredients, leading to a state of sensory alienation where only the “burn” remains.
Evidence Types: Culinary criticism regarding extract-based sauces vs. whole-pepper sauces, The rise of ‘stunt foods’ in fast food marketing, Analogies to ‘shock art’
Rhetorical Approach: Pathos
Est. Words: 250
Counterarguments & Rebuttals
Opposing View: The capsaicin rush is purely about the endorphin high and personal ‘flow state,’ not social performance.
Rebuttal Strategy: Acknowledge the “runner’s high” aspect, but point out that if it were purely private, the multi-million dollar industry of filming the reaction wouldn’t exist. The “high” is now secondary to the “broadcast.”
Est. Words: 100
Opposing View: Spicy food is a cornerstone of many global cultures; calling it a ‘social poison’ is Eurocentric or dismissive of tradition.
Rebuttal Strategy: Distinguish between culinary spice (used for balance, preservation, and flavor in cultures like Thai, Mexican, or Indian) and performative spice (the modern Western “extreme” trend). The latter actually insults the former by reducing complex cuisines to a mere “dare.”
Est. Words: 100
Conclusion Strategy
Reiterate how the “biological honesty” of the pepper has been hijacked by a culture obsessed with performance and “toughness.” Connect the “Spicy Food Semiotic Field” to broader trends in digital culture—where every private bodily experience must be externalized and ranked. Urge a return to “sensory presence” and reclaim the chili pepper as a culinary tool of nuance. End with a final image of a quiet moment of someone enjoying a perfectly balanced, moderately spicy meal—focusing on the flavor, not the camera.
Status: ✅ Complete
Outline Visualization
Prompt:
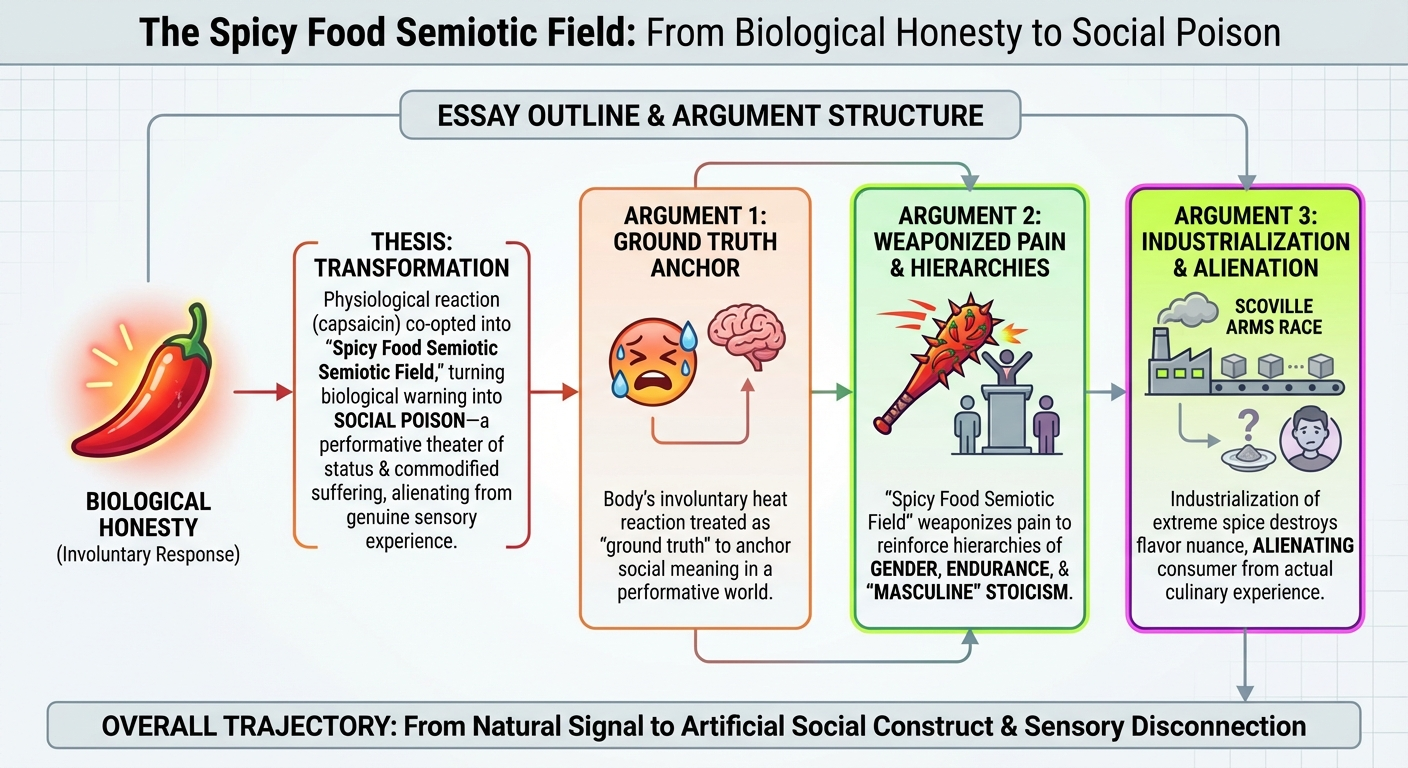
Introduction
A bead of sweat tracks a jagged path through heavy foundation; pupils dilate into obsidian saucers; a panicked, rhythmic gasping replaces the polished cadence of a celebrity interview. This is the high-definition spectacle of the “Hot Ones” era, where the “One Chip Challenge” transforms a private biological crisis into a public digital ritual. We watch, rapt, as the body’s autonomic nervous system revolts in real-time, finding a strange, voyeuristic ecstasy in the sight of a human being pushed to the brink of a physical breakdown.
At its core, this reaction is triggered by capsaicin, a chemical evolved as “directed deterrence”—a molecular lie that tricks our TRPV1 receptors into sensing a thermal fire where none exists. For millennia, humans integrated this “burn” into culinary tapestries for preservation and depth, respecting the pepper as a tool of flavor. Yet, in our hyper-mediated age, the biological response has been decoupled from the dinner plate. We have entered the “Spicy Food Semiotic Field,” a space where the involuntary breakdown of the body—the “unfakeable” honest signal—is no longer a warning, but a prized commodity in a world of curated personas. In this economy of the visceral, the more one suffers, the more “authentic” the content becomes.
However, this shift from seasoning to spectacle carries a hidden cost. While the physiological response to capsaicin is an involuntary “honest signal” of the body, its co-optation into the “Spicy Food Semiotic Field” has transformed a biological warning into a social poison—a performative theater of status, “toughness,” and commodified suffering that ultimately alienates us from genuine sensory experience.
Word Count: 249
Argument 1: We treat the body’s involuntary reaction to heat as a “ground truth” to anchor social meaning in an increasingly performative world.
In an age defined by the hyper-curated digital self, we have come to treat the body’s involuntary reaction to capsaicin as a rare “ground truth”—a biological anchor in a sea of performative artifice. This reliance rests on what evolutionary biologists call “honest signals”: physiological responses, such as the vasodilation of the “capsaicin flush” or spontaneous lacrimation, that are mediated by the TRPV1 heat-sensing receptors and cannot be consciously faked. Unlike a practiced smile or a scripted soundbite, the body’s frantic attempt to cool itself is an unmediated eruption of the physical over the social. We engage in a form of “strategic essentialism,” collectively deciding that the person gasping for air and weeping over a habanero is the “real” individual emerging from behind the mask of etiquette. This is the central conceit of the Hot Ones interview format, which functions less as a culinary show and more as a secular confessional; by subjecting celebrities to escalating levels of commodified pain, the host strips away their PR-managed veneers to reveal a raw, “authentic” character. By designating this involuntary suffering as a semiotic anchor for “grit” or “honesty,” we transform a simple chemical burn into a moral litmus test. However, this fixation on the body’s “truth” ironically deepens our alienation, as we begin to value the spectacle of the reaction more than the sensory reality of the experience itself, setting the stage for the commodification of this very suffering.
Word Count: 236
Argument 1 Image
Prompt:

Argument 2: The “Spicy Food Semiotic Field” weaponizes pain to reinforce hierarchies of gender, endurance, and “masculine” stoicism.
This biological “honesty” is precisely what allows the Spicy Food Semiotic Field to weaponize pain as a metric for social hierarchy, transforming a culinary choice into a performative trial of “frontier masculinity.” In this theater of stoicism, the goal is no longer the appreciation of flavor, but the maintenance of a rigid “poker face” while the body’s TRPV1 receptors scream in agony. Marketing for “extreme” sauces—branded with hyper-masculine imagery of skulls, grenades, and names like The End or Da’ Bomb—recasts the consumer not as a diner, but as a combatant. This mirrors historical rites of passage where enduring physical trauma was a prerequisite for tribal status; today, the “chili-head” subculture utilizes high-Scoville consumption as a “costly signal” of superior constitution. When a social media influencer films themselves consuming a Carolina Reaper without flinching, they are not engaging in a sensory experience; they are participating in a commodified ordeal designed to signal virility and dominance. By equating the suppression of a biological warning with “toughness,” we strip the meal of its communal joy, replacing the pleasure of the palate with a grim, competitive endurance. Ultimately, this transformation alienates the individual from the genuine sensory reality of the food, reducing a complex biological interaction to a hollow trophy of hegemonic stoicism. In doing so, we do not master the spice; we merely use it to mask a deeper hunger for social validation—a hunger that the market is all too eager to feed.
Word Count: 240
Argument 2 Image
Prompt:

Argument 3: The industrialization of extreme spice (the “Scoville Arms Race”) destroys the nuance of flavor, alienating the consumer from the actual culinary experience.
Beyond the performative posturing of the individual, the industrialization of extreme spice—the so-called “Scoville Arms Race”—has effectively lobotomized the culinary arts, replacing the symphony of flavor with a singular, deafening scream of pain. When we prioritize pure capsaicin extracts over the nuanced profile of a whole pepper, we are no longer dining; we are participating in a form of chemical warfare where the palate is the primary casualty. This is the era of “stunt foods,” where products like the “One Chip Challenge” or extract-laden “death sauces” are marketed not as nourishment, but as endurance tests. In this commodified circus, the diner is reduced to a masochistic performer, and the food itself becomes a mere delivery system for a drug-like hit of adrenaline. Much like “shock art” that seeks to offend rather than evoke, these hyper-spicy concoctions achieve a total sensory erasure, blinding the taster to the delicate interplay of acidity, sweetness, and umami. We are left in a state of sensory alienation, where the tongue—once a gateway to cultural history and botanical complexity—is scorched into silence. By turning a biological warning into a commercial spectacle, the industry has stripped the act of eating of its intimacy, leaving us to choke on the ashes of a flavor profile we can no longer feel. This systematic destruction of nuance ensures that we are no longer consumers of culture, but victims of a manufactured crisis, forever chasing a burn that offers no warmth. This erasure of the culinary self paves the way for a deeper disconnection: the total commodification of the body’s survival instincts.
Word Count: 261
Argument 3 Image
Prompt:

Counterarguments & Rebuttals
Critics may claim that the pursuit of heat is a solitary endeavor—a quest for the “capsaicin high” or a meditative flow state triggered by the body’s endorphin response. While the physiological “runner’s high” of spice is undeniable, this defense ignores the modern context. If the experience were truly private, the multi-million dollar industry of “reaction” content and “extreme” marketing would collapse. The internal rush has become secondary to the external broadcast; the high is now merely the fuel for the performance.
Furthermore, some argue that labeling spice as a “social poison” is Eurocentric, dismissing the deep-rooted culinary traditions of the Global South. However, a vital distinction must be made between culinary spice—used for preservation, balance, and flavor in cultures from Mexico to Thailand—and performative spice. The modern Western obsession with “extreme” heat does not honor these traditions; rather, it reduces complex, centuries-old cuisines to a crude “dare.” By stripping spice of its gastronomic context, the performative trend actually insults these cultures, reinforcing the thesis that heat has been weaponized as a tool of status rather than a medium of taste.
Word Count: 180
Counterargument Visualization
Prompt:

Conclusion
The chili pepper was never meant to be a weapon of the ego; it was an evolutionary boundary—a biological “no” that we have perversely rebranded as a social “yes.” By colonizing the TRPV1 receptor with the flags of hyper-masculinity and digital clout, we have transformed an intimate physiological event into a hollow public spectacle. This “Spicy Food Semiotic Field” is a microcosm of our broader cultural malaise: a world where the private sanctity of the body is constantly externalized, ranked, and commodified for the gaze of an invisible audience. When we treat heat as a metric of toughness rather than a component of flavor, we do not just lose the nuance of the pepper; we lose our capacity for genuine sensory presence.
We must dismantle this theater of performative suffering and reclaim the chili as a tool of culinary grace. True epicureanism lies not in the endurance of pain, but in the appreciation of balance. Imagine, instead, a quiet evening where the phone remains dark and the Scoville scale is forgotten. Before you sits a meal where the heat does not scream, but hums—a gentle, radiant warmth that elevates the sweetness of a charred tomato or the citrus notes of a habanero without obliterating the palate. In this silence, the pepper returns to its rightful place: not as a test of the soul, but as a gift to the senses. We owe it to ourselves to stop performing our lives and start tasting them. Eat to feel, not to be seen.
Word Count: 251
Revision Pass 1
Completed revision pass 1 of 2
Revision Pass 2
Completed revision pass 2 of 2
Complete Essay
The Spicy Food Semiotic Field: From Biological Honesty to Social Poison
The Scoville Spectacle: How the Commodification of Pain Alienates the Senses
A bead of sweat carves a salt-stained path through heavy foundation; pupils dilate into obsidian voids; a panicked, rhythmic gasping replaces the polished cadence of a celebrity interview. This is the high-definition spectacle of the “Hot Ones” era, where the “One Chip Challenge” transforms a private biological crisis into a public digital ritual. We watch, rapt, as the body’s autonomic nervous system revolts in real-time, finding a strange, voyeuristic ecstasy in the sight of a human being pushed to the brink of physical collapse.
At its core, this reaction is triggered by capsaicin, a chemical evolved as “directed deterrence”—a molecular lie that tricks our TRPV1 receptors into sensing a thermal fire where none exists. For millennia, humans integrated this “burn” into culinary tapestries for preservation and depth, respecting the pepper as a tool of flavor. Yet, in our hyper-mediated age, the biological response has been decoupled from the dinner plate. We have entered the “Spicy Food Semiotic Field,” a space where the involuntary breakdown of the body—the “unfakeable” honest signal—is no longer a warning, but a prized commodity in a world of curated personas. In this economy of the visceral, the more one suffers, the more “authentic” the content becomes.
However, this shift from seasoning to spectacle carries a hidden cost. While the physiological response to capsaicin is an involuntary “honest signal” of the body, its co-optation into the Spicy Food Semiotic Field has transformed a biological warning into a social poison—a performative theater of status, “toughness,” and commodified suffering that ultimately alienates us from genuine sensory experience.
In an age defined by the hyper-curated digital self, we have come to treat the body’s involuntary reaction to capsaicin as a rare “ground truth”—a biological anchor in a sea of performative artifice. This reliance rests on what evolutionary biologists call “honest signals”: physiological responses, such as the vasodilation of the “capsaicin flush” or spontaneous lacrimation, that are mediated by the nervous system and cannot be consciously faked. Unlike a practiced smile or a scripted soundbite, the body’s frantic attempt to cool itself is an unmediated eruption of the physical over the social.
We engage in a form of “strategic essentialism,” collectively deciding that the person gasping for air and weeping over a habanero is the “real” individual emerging from behind the mask of etiquette. This is the central conceit of the Hot Ones interview format, which functions less as a culinary show and more as a secular confessional. By subjecting celebrities to escalating levels of commodified pain, the host strips away their PR-managed veneers to reveal a raw, “authentic” character. By designating this involuntary suffering as a semiotic anchor for “grit” or “honesty,” we transform a simple chemical burn into a moral litmus test. However, this fixation on the body’s “truth” ironically deepens our alienation; we begin to value the spectacle of the reaction more than the sensory reality of the experience, setting the stage for the total commodification of human distress.
This biological “honesty” is precisely what allows the Spicy Food Semiotic Field to weaponize pain as a metric for social hierarchy, transforming a culinary choice into a performative trial of “frontier masculinity.” In this theater of stoicism, the goal is no longer the appreciation of flavor, but the maintenance of a rigid “poker face” while the body’s receptors scream in agony. Marketing for “extreme” sauces—branded with hyper-masculine imagery of skulls, grenades, and names like The End or Da’ Bomb—recasts the consumer not as a diner, but as a combatant.
This mirrors historical rites of passage where enduring physical trauma was a prerequisite for tribal status; today, the “chili-head” subculture utilizes high-Scoville consumption as a “costly signal” of superior constitution. When a social media influencer films themselves consuming a Carolina Reaper without flinching, they are not engaging in a sensory experience; they are participating in a commodified ordeal designed to signal virility and dominance. By equating the suppression of a biological warning with “toughness,” we strip the meal of its communal joy, replacing the pleasure of the palate with a grim, competitive endurance. Ultimately, this transformation alienates the individual from the genuine sensory reality of the food, reducing a complex biological interaction to a hollow trophy of hegemonic stoicism. In doing so, we do not master the spice; we merely use it to mask a deeper hunger for social validation—a hunger that the market is all too eager to feed.
Beyond the performative posturing of the individual, the industrialization of extreme spice—the so-called “Scoville Arms Race”—has effectively lobotomized the culinary arts, replacing the symphony of flavor with a singular, deafening scream of pain. When we prioritize pure capsaicin extracts over the nuanced profile of a whole pepper, we are no longer dining; we are participating in a form of chemical warfare where the palate is the primary casualty. This is the era of “stunt foods,” where products like the “One Chip Challenge” or extract-laden “death sauces” are marketed not as nourishment, but as endurance tests.
In this commodified circus, the diner is reduced to a masochistic performer, and the food itself becomes a mere delivery system for a drug-like hit of adrenaline. Much like “shock art” that seeks to offend rather than evoke, these hyper-spicy concoctions achieve a total sensory erasure, blinding the taster to the delicate interplay of acidity, sweetness, and umami. We are left in a state of sensory alienation, where the tongue—once a gateway to cultural history and botanical complexity—is scorched into silence. By turning a biological warning into a commercial spectacle, the industry has stripped the act of eating of its intimacy, leaving us to choke on the ashes of a flavor profile we can no longer feel. This systematic destruction of nuance ensures that we are no longer consumers of culture, but victims of a manufactured crisis, forever chasing a burn that offers no warmth. This erasure of the culinary self paves the way for a deeper disconnection: the total commodification of the body’s survival instincts.
Critics may claim that the pursuit of heat is a solitary endeavor—a quest for the “capsaicin high” or a meditative flow state triggered by the body’s endorphin response. While the physiological “runner’s high” of spice is undeniable, this defense ignores the modern context. If the experience were truly private, the multi-million dollar industry of “reaction” content and “extreme” marketing would collapse. The internal rush has become secondary to the external broadcast; the high is now merely the fuel for the performance. We are not chasing enlightenment; we are chasing engagement.
Furthermore, some argue that labeling spice as a “social poison” is Eurocentric, dismissing the deep-rooted culinary traditions of the Global South. However, a vital distinction must be made between culinary spice—used for preservation, balance, and flavor in cultures from Mexico to Thailand—and performative spice. The modern Western obsession with “extreme” heat does not honor these traditions; rather, it reduces complex, centuries-old cuisines to a crude “dare.” By stripping spice of its gastronomic context, the performative trend actually insults these cultures, reinforcing the thesis that heat has been weaponized as a tool of status rather than a medium of taste. The “Scoville Arms Race” is not an homage to global cuisine; it is its caricature.
The chili pepper was never meant to be a weapon of the ego; it was an evolutionary boundary—a biological “no” that we have perversely rebranded as a social “yes.” By colonizing the TRPV1 receptor with the flags of hyper-masculinity and digital clout, we have transformed an intimate physiological event into a hollow public spectacle. This Spicy Food Semiotic Field is a microcosm of our broader cultural malaise: a world where the private sanctity of the body is constantly externalized, ranked, and commodified for the gaze of an invisible audience. When we treat heat as a metric of toughness rather than a component of flavor, we do not just lose the nuance of the pepper; we lose our capacity for genuine sensory presence.
We must dismantle this theater of performative suffering and reclaim the chili as a tool of culinary grace. True epicureanism lies not in the endurance of pain, but in the appreciation of balance. Imagine, instead, a quiet evening where the phone remains dark and the Scoville scale is forgotten. Before you sits a meal where the heat does not scream, but hums—a gentle, radiant warmth that elevates the sweetness of a charred tomato or the citrus notes of a habanero without obliterating the palate. In this silence, the pepper returns to its rightful place: not as a test of the soul, but as a gift to the senses. We owe it to ourselves to stop performing our lives and start tasting them. Eat to feel, not to be seen.
Total Word Count: 1417
Target Word Count: 1200
Completion: 118%
Generation Complete
Total Word Count: 1417
Total Time: 334.627s
Completed: 2026-02-20 21:03:26
Crawler Agent Transcript
Started: 2026-02-20 21:01:45
Search Query: sociology and biology of spicy food capsaicin paradox benign masochism social signaling TRPV1
Direct URLs: N/A
Execution Configuration (click to expand)
1
2
3
4
5
6
7
8
{
"biological_mechanisms" : "Details on TRPV1 receptor polymorphism, 'chili bleaching', and the physiological 'second burn' as evolutionary strategies.",
"psychological_theories" : "Analysis of Paul Rozin's 'benign masochism' and the 'nociceptive-interpretive gap' in the context of safe threats.",
"social_hypotheses" : "Evidence or discussion regarding the 'Performance Hypothesis' (costly signaling/Handicap Principle) and the 'Toughness Hypothesis' (Fundamental Attribution Error) in spice consumption.",
"cultural_semiotics" : "Information on 'authenticity policing', 'culinary orientalism', and the 'Sensory Semiotic Triad' (spicy, sour, bitter) as public vs. private signals.",
"industrial_escalation" : "The impact of the 'Hot-Wing Industrial Complex' (e.g., Hot Ones), Scoville-based marketing, and game-theoretic models like the 'Pooling Equilibrium' or 'Red Queen race' in food trends.",
"philosophical_context" : "References to Elaine Scarry's 'The Body in Pain' or similar works regarding the limits of language and semiotic collapse during intense physical sensation."
}
Crawling Work Details
Error: Failed to get seed items
Stack Trace
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
java.lang.RuntimeException: Search proxy error: HTTP 403
at com.simiacryptus.cognotik.crawl.seed.GoogleProxy$createStrategy$1.getSeedItems(GoogleProxy.kt:61)
at com.simiacryptus.cognotik.plan.tools.online.CrawlerAgentTask.innerRun(CrawlerAgentTask.kt:326)
at com.simiacryptus.cognotik.plan.tools.online.CrawlerAgentTask.run(CrawlerAgentTask.kt:238)
at com.simiacryptus.cognotik.apps.SingleTaskApp.executeTask(SingleTaskApp.kt:127)
at com.simiacryptus.cognotik.apps.SingleTaskApp.startSession$lambda$0(SingleTaskApp.kt:90)
at java.base/java.util.concurrent.Executors$RunnableAdapter.call(Executors.java:572)
at java.base/java.util.concurrent.FutureTask.run$$$capture(FutureTask.java:317)
at java.base/java.util.concurrent.FutureTask.run(FutureTask.java)
at --- Async.Stack.Trace --- (captured by IntelliJ IDEA debugger)
at java.base/java.util.concurrent.FutureTask.<init>(FutureTask.java:151)
at java.base/java.util.concurrent.AbstractExecutorService.newTaskFor(AbstractExecutorService.java:98)
at java.base/java.util.concurrent.AbstractExecutorService.submit(AbstractExecutorService.java:122)
at com.simiacryptus.cognotik.util.ImmediateExecutorService.submit(ImmediateExecutorService.kt:77)
at com.simiacryptus.cognotik.apps.SingleTaskApp.startSession(SingleTaskApp.kt:90)
at com.simiacryptus.cognotik.apps.SingleTaskApp.newSession(SingleTaskApp.kt:58)
at com.simiacryptus.cognotik.util.UnifiedHarness$runTask$singleTaskApp$1.newSession(UnifiedHarness.kt:274)
at com.simiacryptus.cognotik.util.UnifiedHarness.runTask(UnifiedHarness.kt:290)
at com.simiacryptus.cognotik.util.DocProcessor.runAll$lambda$3$0(DocProcessor.kt:788)
at com.simiacryptus.cognotik.util.FixedConcurrencyProcessor.executeTask$lambda$0(FixedConcurrencyProcessor.kt:97)
at java.base/java.util.concurrent.CompletableFuture$AsyncSupply.run$$$capture(CompletableFuture.java:1768)
at java.base/java.util.concurrent.CompletableFuture$AsyncSupply.run(CompletableFuture.java)
at --- Async.Stack.Trace --- (captured by IntelliJ IDEA debugger)
at java.base/java.util.concurrent.CompletableFuture$AsyncSupply.<init>(CompletableFuture.java:1754)
at java.base/java.util.concurrent.CompletableFuture.asyncSupplyStage(CompletableFuture.java:1782)
at java.base/java.util.concurrent.CompletableFuture.supplyAsync(CompletableFuture.java:2005)
at com.simiacryptus.cognotik.util.FixedConcurrencyProcessor.executeTask(FixedConcurrencyProcessor.kt:91)
at com.simiacryptus.cognotik.util.FixedConcurrencyProcessor.tryExecuteTask(FixedConcurrencyProcessor.kt:79)
at com.simiacryptus.cognotik.util.FixedConcurrencyProcessor.processNextWaitingTask(FixedConcurrencyProcessor.kt:129)
at com.simiacryptus.cognotik.util.FixedConcurrencyProcessor.executeTask$lambda$1(FixedConcurrencyProcessor.kt:118)
at com.simiacryptus.cognotik.util.FixedConcurrencyProcessor.executeTask$lambda$2(FixedConcurrencyProcessor.kt:104)
at java.base/java.util.concurrent.CompletableFuture.uniWhenComplete(CompletableFuture.java:863)
at java.base/java.util.concurrent.CompletableFuture$UniWhenComplete.tryFire$$$capture(CompletableFuture.java:841)
at java.base/java.util.concurrent.CompletableFuture$UniWhenComplete.tryFire(CompletableFuture.java)
at --- Async.Stack.Trace --- (captured by IntelliJ IDEA debugger)
at java.base/java.util.concurrent.CompletableFuture$UniWhenComplete.<init>(CompletableFuture.java:834)
at java.base/java.util.concurrent.CompletableFuture.uniWhenCompleteStage(CompletableFuture.java:885)
at java.base/java.util.concurrent.CompletableFuture.whenComplete(CompletableFuture.java:2357)
at com.simiacryptus.cognotik.util.FixedConcurrencyProcessor.executeTask(FixedConcurrencyProcessor.kt:104)
at com.simiacryptus.cognotik.util.FixedConcurrencyProcessor.tryExecuteTask(FixedConcurrencyProcessor.kt:79)
at com.simiacryptus.cognotik.util.FixedConcurrencyProcessor.processNextWaitingTask(FixedConcurrencyProcessor.kt:129)
at com.simiacryptus.cognotik.util.FixedConcurrencyProcessor.executeTask$lambda$1(FixedConcurrencyProcessor.kt:118)
at com.simiacryptus.cognotik.util.FixedConcurrencyProcessor.executeTask$lambda$2(FixedConcurrencyProcessor.kt:104)
at java.base/java.util.concurrent.CompletableFuture.uniWhenComplete(CompletableFuture.java:863)
at java.base/java.util.concurrent.CompletableFuture$UniWhenComplete.tryFire$$$capture(CompletableFuture.java:841)
at java.base/java.util.concurrent.CompletableFuture$UniWhenComplete.tryFire(CompletableFuture.java)
at --- Async.Stack.Trace --- (captured by IntelliJ IDEA debugger)
at java.base/java.util.concurrent.CompletableFuture$UniWhenComplete.<init>(CompletableFuture.java:834)
at java.base/java.util.concurrent.CompletableFuture.uniWhenCompleteStage(CompletableFuture.java:885)
at java.base/java.util.concurrent.CompletableFuture.whenComplete(CompletableFuture.java:2357)
at com.simiacryptus.cognotik.util.FixedConcurrencyProcessor.executeTask(FixedConcurrencyProcessor.kt:104)
at com.simiacryptus.cognotik.util.FixedConcurrencyProcessor.tryExecuteTask(FixedConcurrencyProcessor.kt:79)
at com.simiacryptus.cognotik.util.FixedConcurrencyProcessor.processNextWaitingTask(FixedConcurrencyProcessor.kt:129)
at com.simiacryptus.cognotik.util.FixedConcurrencyProcessor.executeTask$lambda$1(FixedConcurrencyProcessor.kt:118)
at com.simiacryptus.cognotik.util.FixedConcurrencyProcessor.executeTask$lambda$2(FixedConcurrencyProcessor.kt:104)
at java.base/java.util.concurrent.CompletableFuture.uniWhenComplete(CompletableFuture.java:863)
at java.base/java.util.concurrent.CompletableFuture$UniWhenComplete.tryFire$$$capture(CompletableFuture.java:841)
at java.base/java.util.concurrent.CompletableFuture$UniWhenComplete.tryFire(CompletableFuture.java)
at --- Async.Stack.Trace --- (captured by IntelliJ IDEA debugger)
at java.base/java.util.concurrent.CompletableFuture$UniWhenComplete.<init>(CompletableFuture.java:834)
at java.base/java.util.concurrent.CompletableFuture.uniWhenCompleteStage(CompletableFuture.java:885)
at java.base/java.util.concurrent.CompletableFuture.whenComplete(CompletableFuture.java:2357)
at com.simiacryptus.cognotik.util.FixedConcurrencyProcessor.executeTask(FixedConcurrencyProcessor.kt:104)
at com.simiacryptus.cognotik.util.FixedConcurrencyProcessor.tryExecuteTask(FixedConcurrencyProcessor.kt:79)
at com.simiacryptus.cognotik.util.FixedConcurrencyProcessor.processNextWaitingTask(FixedConcurrencyProcessor.kt:129)
at com.simiacryptus.cognotik.util.FixedConcurrencyProcessor.executeTask$lambda$1(FixedConcurrencyProcessor.kt:118)
at com.simiacryptus.cognotik.util.FixedConcurrencyProcessor.executeTask$lambda$2(FixedConcurrencyProcessor.kt:104)
at java.base/java.util.concurrent.CompletableFuture.uniWhenComplete(CompletableFuture.java:863)
at java.base/java.util.concurrent.CompletableFuture$UniWhenComplete.tryFire$$$capture(CompletableFuture.java:841)
at java.base/java.util.concurrent.CompletableFuture$UniWhenComplete.tryFire(CompletableFuture.java)
at --- Async.Stack.Trace --- (captured by IntelliJ IDEA debugger)
at java.base/java.util.concurrent.CompletableFuture$UniWhenComplete.<init>(CompletableFuture.java:834)
at java.base/java.util.concurrent.CompletableFuture.uniWhenCompleteStage(CompletableFuture.java:885)
at java.base/java.util.concurrent.CompletableFuture.whenComplete(CompletableFuture.java:2357)
at com.simiacryptus.cognotik.util.FixedConcurrencyProcessor.executeTask(FixedConcurrencyProcessor.kt:104)
at com.simiacryptus.cognotik.util.FixedConcurrencyProcessor.tryExecuteTask(FixedConcurrencyProcessor.kt:79)
at com.simiacryptus.cognotik.util.FixedConcurrencyProcessor.processNextWaitingTask(FixedConcurrencyProcessor.kt:129)
at com.simiacryptus.cognotik.util.FixedConcurrencyProcessor.executeTask$lambda$1(FixedConcurrencyProcessor.kt:118)
at com.simiacryptus.cognotik.util.FixedConcurrencyProcessor.executeTask$lambda$2(FixedConcurrencyProcessor.kt:104)
at java.base/java.util.concurrent.CompletableFuture.uniWhenComplete(CompletableFuture.java:863)
at java.base/java.util.concurrent.CompletableFuture$UniWhenComplete.tryFire$$$capture(CompletableFuture.java:841)
at java.base/java.util.concurrent.CompletableFuture$UniWhenComplete.tryFire(CompletableFuture.java)
at --- Async.Stack.Trace --- (captured by IntelliJ IDEA debugger)
at java.base/java.util.concurrent.CompletableFuture$UniWhenComplete.<init>(CompletableFuture.java:834)
at java.base/java.util.concurrent.CompletableFuture.uniWhenCompleteStage(CompletableFuture.java:885)
at java.base/java.util.concurrent.CompletableFuture.whenComplete(CompletableFuture.java:2357)
at com.simiacryptus.cognotik.util.FixedConcurrencyProcessor.executeTask(FixedConcurrencyProcessor.kt:104)
at com.simiacryptus.cognotik.util.FixedConcurrencyProcessor.tryExecuteTask(FixedConcurrencyProcessor.kt:79)
at com.simiacryptus.cognotik.util.FixedConcurrencyProcessor.submit(FixedConcurrencyProcessor.kt:53)
at com.simiacryptus.cognotik.util.DocProcessor.runAll(DocProcessor.kt:772)
at com.simiacryptus.cognotik.util.DocProcessor.runAll$default(DocProcessor.kt:765)
at cognotik.actions.task.DocProcessorAction$handle$1$1.run(DocProcessorAction.kt:172)
at com.intellij.openapi.progress.impl.CoreProgressManager.startTask(CoreProgressManager.java:491)
at com.intellij.openapi.progress.impl.ProgressManagerImpl.startTask(ProgressManagerImpl.java:133)
at com.intellij.openapi.progress.impl.CoreProgressManager.lambda$runProcessWithProgressAsynchronously$7(CoreProgressManager.java:542)
at com.intellij.openapi.progress.impl.ProgressRunner.lambda$submit$4(ProgressRunner.java:249)
at com.intellij.openapi.progress.ProgressManager.lambda$runProcess$0(ProgressManager.java:98)
at com.intellij.openapi.progress.impl.CoreProgressManager.lambda$runProcess$1(CoreProgressManager.java:223)
at com.intellij.platform.diagnostic.telemetry.helpers.TraceKt.use(trace.kt:45)
at com.intellij.openapi.progress.impl.CoreProgressManager.lambda$runProcess$2(CoreProgressManager.java:222)
at com.intellij.openapi.progress.impl.CoreProgressManager.lambda$executeProcessUnderProgress$14(CoreProgressManager.java:674)
at com.intellij.openapi.progress.impl.CoreProgressManager.registerIndicatorAndRun(CoreProgressManager.java:749)
at com.intellij.openapi.progress.impl.CoreProgressManager.computeUnderProgress(CoreProgressManager.java:705)
at com.intellij.openapi.progress.impl.CoreProgressManager.executeProcessUnderProgress(CoreProgressManager.java:673)
at com.intellij.openapi.progress.impl.ProgressManagerImpl.executeProcessUnderProgress(ProgressManagerImpl.java:79)
at com.intellij.openapi.progress.impl.CoreProgressManager.runProcess(CoreProgressManager.java:203)
at com.intellij.openapi.progress.ProgressManager.runProcess(ProgressManager.java:98)
at com.intellij.openapi.progress.impl.ProgressRunner.lambda$submit$5(ProgressRunner.java:249)
at com.intellij.openapi.progress.impl.ProgressRunner$ProgressRunnable.run$$$capture(ProgressRunner.java:502)
at com.intellij.openapi.progress.impl.ProgressRunner$ProgressRunnable.run(ProgressRunner.java)
at --- Async.Stack.Trace --- (captured by IntelliJ IDEA debugger)
at com.intellij.openapi.progress.impl.ProgressRunner$ProgressRunnable.<init>(ProgressRunner.java:492)
at com.intellij.openapi.progress.impl.ProgressRunner.lambda$launchTask$20(ProgressRunner.java:461)
at java.base/java.util.concurrent.CompletableFuture.uniWhenComplete(CompletableFuture.java:863)
at java.base/java.util.concurrent.CompletableFuture.uniWhenCompleteStage(CompletableFuture.java:887)
at java.base/java.util.concurrent.CompletableFuture.whenComplete(CompletableFuture.java:2357)
at com.intellij.openapi.progress.impl.ProgressRunner.launchTask(ProgressRunner.java:456)
at com.intellij.openapi.progress.impl.ProgressRunner.execFromEDT(ProgressRunner.java:303)
at com.intellij.openapi.progress.impl.ProgressRunner.submit(ProgressRunner.java:252)
at com.intellij.openapi.progress.impl.CoreProgressManager.runProcessWithProgressAsynchronously(CoreProgressManager.java:550)
at com.intellij.openapi.progress.impl.CoreProgressManager.runProcessWithProgressAsynchronously(CoreProgressManager.java:484)
at com.intellij.openapi.progress.impl.CoreProgressManager.runProcessWithProgressAsynchronously(CoreProgressManager.java:476)
at com.intellij.openapi.progress.impl.CoreProgressManager.runAsynchronously(CoreProgressManager.java:453)
at com.intellij.openapi.progress.impl.CoreProgressManager.run(CoreProgressManager.java:436)
at cognotik.actions.task.DocProcessorAction.handle$lambda$3(DocProcessorAction.kt:161)
at com.intellij.openapi.application.impl.AnyThreadWriteThreadingSupport.runIntendedWriteActionOnCurrentThread$lambda$2(AnyThreadWriteThreadingSupport.kt:217)
at com.intellij.openapi.application.impl.AnyThreadWriteThreadingSupport.runWriteIntentReadAction(AnyThreadWriteThreadingSupport.kt:128)
at com.intellij.openapi.application.impl.AnyThreadWriteThreadingSupport.runIntendedWriteActionOnCurrentThread(AnyThreadWriteThreadingSupport.kt:216)
at com.intellij.openapi.application.impl.ApplicationImpl.runIntendedWriteActionOnCurrentThread(ApplicationImpl.java:842)
at com.intellij.openapi.application.impl.ApplicationImpl.invokeAndWait(ApplicationImpl.java:395)
at com.intellij.openapi.application.impl.ApplicationImpl.invokeAndWait(ApplicationImpl.java:446)
at cognotik.actions.task.DocProcessorAction.handle(DocProcessorAction.kt:155)
at cognotik.actions.BaseAction.actionPerformed(BaseAction.kt:55)
at com.intellij.openapi.actionSystem.ex.ActionUtil.doPerformActionOrShowPopup(ActionUtil.kt:374)
at com.intellij.openapi.actionSystem.ex.ActionUtil.performActionDumbAwareWithCallbacks$lambda$7(ActionUtil.kt:343)
at com.intellij.openapi.actionSystem.impl.ActionManagerImpl.performWithActionCallbacks(ActionManagerImpl.kt:1173)
at com.intellij.openapi.actionSystem.ex.ActionUtil.performActionDumbAwareWithCallbacks(ActionUtil.kt:342)
at com.intellij.openapi.actionSystem.impl.ActionMenuItem.performAction$lambda$5(ActionMenuItem.kt:273)
at com.intellij.openapi.wm.impl.FocusManagerImpl.runOnOwnContext(FocusManagerImpl.java:231)
at com.intellij.openapi.actionSystem.impl.ActionMenuItem.performAction(ActionMenuItem.kt:265)
at com.intellij.openapi.actionSystem.impl.ActionMenuItem._init_$lambda$0(ActionMenuItem.kt:72)
at java.desktop/javax.swing.AbstractButton.fireActionPerformed(AbstractButton.java:1972)
at com.intellij.openapi.actionSystem.impl.ActionMenuItem.fireActionPerformed$lambda$4(ActionMenuItem.kt:103)
at com.intellij.openapi.application.TransactionGuardImpl.performActivity(TransactionGuardImpl.java:109)
at com.intellij.openapi.application.TransactionGuardImpl.performUserActivity(TransactionGuardImpl.java:98)
at com.intellij.openapi.actionSystem.impl.ActionMenuItem.fireActionPerformed(ActionMenuItem.kt:102)
at com.intellij.ui.plaf.beg.BegMenuItemUI.doClick(BegMenuItemUI.java:521)
at com.intellij.ui.plaf.beg.BegMenuItemUI$MyMouseInputHandler.mouseReleased(BegMenuItemUI.java:554)
at java.desktop/java.awt.Component.processMouseEvent(Component.java:6662)
at java.desktop/javax.swing.JComponent.processMouseEvent(JComponent.java:3394)
at java.desktop/java.awt.Component.processEvent(Component.java:6427)
at java.desktop/java.awt.Container.processEvent(Container.java:2266)
at java.desktop/java.awt.Component.dispatchEventImpl(Component.java:5032)
at java.desktop/java.awt.Container.dispatchEventImpl(Container.java:2324)
at java.desktop/java.awt.Component.dispatchEvent(Component.java:4860)
at java.desktop/java.awt.LightweightDispatcher.retargetMouseEvent(Container.java:4963)
at java.desktop/java.awt.LightweightDispatcher.processMouseEvent(Container.java:4577)
at java.desktop/java.awt.LightweightDispatcher.dispatchEvent(Container.java:4518)
at java.desktop/java.awt.Container.dispatchEventImpl(Container.java:2310)
at java.desktop/java.awt.Window.dispatchEventImpl(Window.java:2810)
at java.desktop/java.awt.Component.dispatchEvent(Component.java:4860)
at java.desktop/java.awt.EventQueue.dispatchEventImpl(EventQueue.java:783)
at java.desktop/java.awt.EventQueue$4.run(EventQueue.java:728)
at java.desktop/java.awt.EventQueue$4.run(EventQueue.java:722)
at java.base/java.security.AccessController.doPrivileged(AccessController.java:400)
at java.base/java.security.ProtectionDomain$JavaSecurityAccessImpl.doIntersectionPrivilege(ProtectionDomain.java:87)
at java.base/java.security.ProtectionDomain$JavaSecurityAccessImpl.doIntersectionPrivilege(ProtectionDomain.java:98)
at java.desktop/java.awt.EventQueue$5.run(EventQueue.java:755)
at java.desktop/java.awt.EventQueue$5.run(EventQueue.java:753)
at java.base/java.security.AccessController.doPrivileged(AccessController.java:400)
at java.base/java.security.ProtectionDomain$JavaSecurityAccessImpl.doIntersectionPrivilege(ProtectionDomain.java:87)
at java.desktop/java.awt.EventQueue.dispatchEvent(EventQueue.java:752)
at com.intellij.ide.IdeEventQueue.defaultDispatchEvent(IdeEventQueue.kt:675)
at com.intellij.ide.IdeEventQueue.dispatchMouseEvent(IdeEventQueue.kt:621)
at com.intellij.ide.IdeEventQueue._dispatchEvent$lambda$21(IdeEventQueue.kt:564)
at com.intellij.openapi.application.impl.AnyThreadWriteThreadingSupport.runWriteIntentReadAction(AnyThreadWriteThreadingSupport.kt:128)
at com.intellij.ide.IdeEventQueue._dispatchEvent(IdeEventQueue.kt:564)
at com.intellij.ide.IdeEventQueue.dispatchEvent$lambda$18$lambda$17$lambda$16$lambda$15(IdeEventQueue.kt:355)
at com.intellij.openapi.progress.impl.CoreProgressManager.computePrioritized(CoreProgressManager.java:857)
at com.intellij.ide.IdeEventQueue.dispatchEvent$lambda$18$lambda$17$lambda$16(IdeEventQueue.kt:354)
at com.intellij.ide.IdeEventQueueKt.performActivity$lambda$2$lambda$1(IdeEventQueue.kt:1045)
at com.intellij.openapi.application.WriteIntentReadAction.lambda$run$0(WriteIntentReadAction.java:24)
at com.intellij.openapi.application.impl.AnyThreadWriteThreadingSupport.runWriteIntentReadAction(AnyThreadWriteThreadingSupport.kt:128)
at com.intellij.openapi.application.impl.ApplicationImpl.runWriteIntentReadAction(ApplicationImpl.java:916)
at com.intellij.openapi.application.WriteIntentReadAction.compute(WriteIntentReadAction.java:55)
at com.intellij.openapi.application.WriteIntentReadAction.run(WriteIntentReadAction.java:23)
at com.intellij.ide.IdeEventQueueKt.performActivity$lambda$2(IdeEventQueue.kt:1045)
at com.intellij.ide.IdeEventQueueKt.performActivity$lambda$3(IdeEventQueue.kt:1054)
at com.intellij.openapi.application.TransactionGuardImpl.performActivity(TransactionGuardImpl.java:117)
at com.intellij.ide.IdeEventQueueKt.performActivity(IdeEventQueue.kt:1054)
at com.intellij.ide.IdeEventQueue.dispatchEvent$lambda$18(IdeEventQueue.kt:349)
at com.intellij.ide.IdeEventQueue.dispatchEvent(IdeEventQueue.kt:395)
at java.desktop/java.awt.EventDispatchThread.pumpOneEventForFilters(EventDispatchThread.java:207)
at java.desktop/java.awt.EventDispatchThread.pumpEventsForFilter(EventDispatchThread.java:128)
at java.desktop/java.awt.EventDispatchThread.pumpEventsForHierarchy(EventDispatchThread.java:117)
at java.desktop/java.awt.EventDispatchThread.pumpEvents(EventDispatchThread.java:113)
at java.desktop/java.awt.EventDispatchThread.pumpEvents(EventDispatchThread.java:105)
at java.desktop/java.awt.EventDispatchThread.run(EventDispatchThread.java:92)